microRNA
Diagram of micro RNA (miRNA) action with mRNA

Examples of miRNA stem-loops, with the mature miRNAs shown in red
A microRNA (abbreviated miRNA) is a small non-coding RNA molecule (containing about 22 nucleotides) found in plants, animals and some viruses, that functions in RNA silencing and post-transcriptional regulation of gene expression.[1][2][3] miRNAs function via base-pairing with complementary sequences within mRNA molecules.[4] As a result, these mRNA molecules are silenced, by one or more of the following processes: (1) Cleavage of the mRNA strand into two pieces, (2) Destabilization of the mRNA through shortening of its poly(A) tail, and (3) Less efficient translation of the mRNA into proteins by ribosomes.[4][5]
miRNAs resemble the small interfering RNAs (siRNAs) of the RNA interference (RNAi) pathway, except miRNAs derive from regions of RNA transcripts that fold back on themselves to form short hairpins, whereas siRNAs derive from longer regions of double-stranded RNA.[2] The human genome may encode over 1900 miRNAs,[6] although more recent analysis indicates that the number is closer to 600.[7]
miRNAs are abundant in many mammalian cell types[8][9] and appear to target about 60% of the genes of humans and other mammals.[10][11] Many miRNAs are evolutionarily conserved, which implies that they have important biological functions.[7][3] For example, 90 families of miRNAs have been conserved since at least the common ancestor of mammals and fish, and most of these conserved miRNAs have important functions, as shown by studies in which genes for one or more members of a family have been knocked out in mouse.[3]
.mw-parser-output .toclimit-2 .toclevel-1 ul,.mw-parser-output .toclimit-3 .toclevel-2 ul,.mw-parser-output .toclimit-4 .toclevel-3 ul,.mw-parser-output .toclimit-5 .toclevel-4 ul,.mw-parser-output .toclimit-6 .toclevel-5 ul,.mw-parser-output .toclimit-7 .toclevel-6 uldisplay:none
Contents
1 History
2 Nomenclature
3 Targets
4 Biogenesis
4.1 Transcription
4.2 Nuclear processing
4.3 Nuclear export
4.4 Cytoplasmic processing
4.5 Biogenesis in plants
5 RNA-induced silencing complex
5.1 Mode of silencing and regulatory loops
6 Turnover
7 Cellular functions
8 Evolution
9 Experimental detection and manipulation
10 Disease
10.1 Inherited diseases
10.2 Cancer
10.3 DNA repair and cancer
10.4 Heart disease
10.4.1 miRNA-712
10.4.2 Gene origin
10.4.3 Mechanism
10.4.4 Human homolog microRNA-205
10.5 Kidney disease
10.6 Nervous system
10.6.1 Alcoholism
10.7 Obesity
10.8 Hemostasis
11 Non-coding RNAs
12 Viruses
13 Target prediction
14 See also
15 References
16 Further reading
17 External links
History
The first miRNA was discovered in the early 1990s.[12] However, miRNAs were not recognized as a distinct class of biological regulators until the early 2000s.[13][14][15][16][17] miRNA research revealed different sets of miRNAs expressed in different cell types and tissues[9][18] and multiple roles for miRNAs in plant and animal development and in many other biological processes.[19][20][21][22][23][24][25][26] Aberrant miRNA expression are implicated in disease states. MiRNA-based therapies are under investigation.[27][28][29][30]
The first miRNA was discovered in 1993 by a group led by Ambros and including Lee and Feinbaum. However, additional insight into its mode of action required simultaneously published work by Ruvkun's team, including Wightman and Ha.[12][31] These groups published back-to-back papers on the lin-4 gene, which was known to control the timing of C. elegans larval development by repressing the lin-14 gene. When Lee et al. isolated the lin-4 miRNA, they found that instead of producing an mRNA encoding a protein, it produced short non-coding RNAs, one of which was a ~22-nucleotide RNA that contained sequences partially complementary to multiple sequences in the 3' UTR of the lin-14 mRNA.[12] This complementarity was proposed to inhibit the translation of the lin-14 mRNA into the LIN-14 protein. At the time, the lin-4 small RNA was thought to be a nematode idiosyncrasy.
In 2000, a second small RNA was characterized: let-7 RNA, which represses lin-41 to promote a later developmental transition in C. elegans.[13] The let-7 RNA was found to be conserved in many species, leading to the suggestion that let-7 RNA and additional "small temporal RNAs" might regulate the timing of development in diverse animals, including humans.[14]
A year later, the lin-4 and let-7 RNAs were found to be part of a large class of small RNAs present in C. elegans, Drosophila and human cells.[15][16][17] The many RNAs of this class resembled the lin-4 and let-7 RNAs, except their expression patterns were usually inconsistent with a role in regulating the timing of development. This suggested that most might function in other types of regulatory pathways. At this point, researchers started using the term "microRNA" to refer to this class of small regulatory RNAs.[15][16][17]
The first human disease associated with deregulation of miRNAs was chronic lymphocytic leukemia.
Nomenclature
Under a standard nomenclature system, names are assigned to experimentally confirmed miRNAs before publication.[32][33] The prefix "miR" is followed by a dash and a number, the latter often indicating order of naming. For example, miR-124 was named and likely discovered prior to miR-456. A capitalized "miR-" refers to the mature form of the miRNA, while the uncapitalized "mir-" refers to the pre-miRNA and the pri-miRNA, and "MIR" refers to the gene that encodes them.[34]
miRNAs with nearly identical sequences except for one or two nucleotides are annotated with an additional lower case letter. For example, miR-124a is closely related to miR-124b.
Pre-miRNAs, pri-miRNAs and genes that lead to 100% identical mature miRNAs but that are located at different places in the genome are indicated with an additional dash-number suffix. For example, the pre-miRNAs hsa-mir-194-1 and hsa-mir-194-2 lead to an identical mature miRNA (hsa-miR-194) but are from genes located in different genome regions.
Species of origin is designated with a three-letter prefix, e.g., hsa-miR-124 is a human (Homo sapiens) miRNA and oar-miR-124 is a sheep (Ovis aries) miRNA. Other common prefixes include 'v' for viral (miRNA encoded by a viral genome) and 'd' for Drosophila miRNA (a fruit fly commonly studied in genetic research).
When two mature microRNAs originate from opposite arms of the same pre-miRNA and are found in roughly similar amounts, they are denoted with a -3p or -5p suffix. (In the past, this distinction was also made with 's' (sense) and 'as' (antisense)). However, the mature microRNA found from one arm of the hairpin is usually much more abundant than that found from the other arm,[2] in which case, an asterisk following the name indicates the mature species found at low levels from the opposite arm of a hairpin. For example, miR-124 and miR-124* share a pre-miRNA hairpin, but much more miR-124 is found in the cell.
Targets
Plant miRNAs usually have near-perfect pairing with their mRNA targets, which induces gene repression through cleavage of the target transcripts.[19] In contrast, animal miRNAs are able to recognize their target mRNAs by using as little as 6–8 nucleotides (the seed region) at the 5' end of the miRNA,[10][35][36] which is not enough pairing to induce cleavage of the target mRNAs.[4] Combinatorial regulation is a feature of miRNA regulation in animals.[4][37] A given miRNA may have hundreds of different mRNA targets, and a given target might be regulated by multiple miRNAs.[11][38]
Estimates of the average number of unique messenger RNAs that are targets for repression by a typical miRNA vary, depending on the estimation method,[39] but multiple approaches show that mammalian miRNAs can have many unique targets. For example, an analysis of the miRNAs highly conserved in vertebrates shows that each has, on average, roughly 400 conserved targets.[11] Likewise, experiments show that a single miRNA species can reduce the stability of hundreds of unique messenger RNAs.[40] Other experiments show that a single miRNA species may repress the production of hundreds of proteins, but that this repression often is relatively mild (much less than 2-fold).[41][42] The first human disease associated with deregulation of miRNAs was chronic lymphocytic leukemia. Other B cell malignancies followed.
Biogenesis

As many as 40% of miRNA genes may lie in the introns or even exons of other genes.[43] These are usually, though not exclusively, found in a sense orientation,[44][45] and thus usually are regulated together with their host genes.[43][46][47]
The DNA template is not the final word on mature miRNA production: 6% of human miRNAs show RNA editing (IsomiRs), the site-specific modification of RNA sequences to yield products different from those encoded by their DNA. This increases the diversity and scope of miRNA action beyond that implicated from the genome alone.
Transcription
miRNA genes are usually transcribed by RNA polymerase II (Pol II).[48][49] The polymerase often binds to a promoter found near the DNA sequence, encoding what will become the hairpin loop of the pre-miRNA. The resulting transcript is capped with a specially modified nucleotide at the 5' end, polyadenylated with multiple adenosines (a poly(A) tail),[48][44] and spliced. Animal miRNAs are initially transcribed as part of one arm of an ∼80 nucleotide RNA stem-loop that in turn forms part of a several hundred nucleotide-long miRNA precursor termed a primary miRNA (pri-miRNA).[48][44] When a stem-loop precursor is found in the 3' UTR, a transcript may serve as a pri-miRNA and a mRNA.[44]RNA polymerase III (Pol III) transcribes some miRNAs, especially those with upstream Alu sequences, transfer RNAs (tRNAs), and mammalian wide interspersed repeat (MWIR) promoter units.[50]
Nuclear processing

A crystal structure of the human Drosha protein in complex with the C-terminal helices of two DGCR8 molecules (green). Drosha consists of two ribonuclease III domains (blue and orange); a double-stranded RNA binding domain (yellow); and a connector/platform domain (gray) containing two bound zinc ion (spheres). From PDB: 5B16.
A single pri-miRNA may contain from one to six miRNA precursors. These hairpin loop structures are composed of about 70 nucleotides each. Each hairpin is flanked by sequences necessary for efficient processing.
The double-stranded RNA (dsRNA) structure of the hairpins in a pri-miRNA is recognized by a nuclear protein known as DiGeorge Syndrome Critical Region 8 (DGCR8 or "Pasha" in invertebrates), named for its association with DiGeorge Syndrome. DGCR8 associates with the enzyme Drosha, a protein that cuts RNA, to form the Microprocessor complex.[51][52] In this complex, DGCR8 orients the catalytic RNase III domain of Drosha to liberate hairpins from pri-miRNAs by cleaving RNA about eleven nucleotides from the hairpin base (one helical dsRNA turn into the stem).[53][54] The product resulting has a two-nucleotide overhang at its 3' end; it has 3' hydroxyl and 5' phosphate groups. It is often termed as a pre-miRNA (precursor-miRNA). Sequence motifs downstream of the pre-miRNA that are important for efficient processing have been identified.[55][56][57]
Pre-miRNAs that are spliced directly out of introns, bypassing the Microprocessor complex, are known as "Mirtrons." Originally thought to exist only in Drosophila and C. elegans, mirtrons have now been found in mammals.[58]
As many as 16% of pre-miRNAs may be altered through nuclear RNA editing.[59][60][61] Most commonly, enzymes known as adenosine deaminases acting on RNA (ADARs) catalyze adenosine to inosine (A to I) transitions. RNA editing can halt nuclear processing (for example, of pri-miR-142, leading to degradation by the ribonuclease Tudor-SN) and alter downstream processes including cytoplasmic miRNA processing and target specificity (e.g., by changing the seed region of miR-376 in the central nervous system).[59]
Nuclear export

The human exportin-5 protein (red) in complex with Ran-GTP (yellow) and a pre-microRNA (green), showing two-nucleotide overhang recognition element (orange). From PDB: 3A6P.
Pre-miRNA hairpins are exported from the nucleus in a process involving the nucleocytoplasmic shuttler Exportin-5. This protein, a member of the karyopherin family, recognizes a two-nucleotide overhang left by the RNase III enzyme Drosha at the 3' end of the pre-miRNA hairpin. Exportin-5-mediated transport to the cytoplasm is energy-dependent, using GTP bound to the Ran protein.[62]
Cytoplasmic processing
In the cytoplasm, the pre-miRNA hairpin is cleaved by the RNase III enzyme Dicer.[63] This endoribonuclease interacts with 5' and 3' ends of the hairpin[64] and cuts away the loop joining the 3' and 5' arms, yielding an imperfect miRNA:miRNA* duplex about 22 nucleotides in length.[63] Overall hairpin length and loop size influence the efficiency of Dicer processing. The imperfect nature of the miRNA:miRNA* pairing also affects cleavage.[63][65] Some of the G-rich pre-miRNAs can potentially adopt the G-quadruplex structure as an alternative to the canonical stem-loop structure. For example, human pre-miRNA 92b adopts a G-quadruplex structure which is resistant to the Dicer mediated cleavage in the cytoplasm.[66] Although either strand of the duplex may potentially act as a functional miRNA, only one strand is usually incorporated into the RNA-induced silencing complex (RISC) where the miRNA and its mRNA target interact.
While the majority of miRNAs are located within the cell, some miRNAs, commonly known as circulating miRNAs or extracellular miRNAs, have also been found in extracellular environment, including various biological fluids and cell culture media.[67][68]
Biogenesis in plants
miRNA biogenesis in plants differs from animal biogenesis mainly in the steps of nuclear processing and export. Instead of being cleaved by two different enzymes, once inside and once outside the nucleus, both cleavages of the plant miRNA are performed by a Dicer homolog, called Dicer-like1 (DL1). DL1 is expressed only in the nucleus of plant cells, which indicates that both reactions take place inside the nucleus. Before plant miRNA:miRNA* duplexes are transported out of the nucleus, its 3' overhangs are methylated by a RNA methyltransferaseprotein called Hua-Enhancer1 (HEN1). The duplex is then transported out of the nucleus to the cytoplasm by a protein called Hasty (HST), an Exportin 5 homolog, where they disassemble and the mature miRNA is incorporated into the RISC.[69]
RNA-induced silencing complex
The mature miRNA is part of an active RNA-induced silencing complex (RISC) containing Dicer and many associated proteins.[70] RISC is also known as a microRNA ribonucleoprotein complex (miRNP);[71] A RISC with incorporated miRNA is sometimes referred to as a "miRISC."
Dicer processing of the pre-miRNA is thought to be coupled with unwinding of the duplex. Generally, only one strand is incorporated into the miRISC, selected on the basis of its thermodynamic instability and weaker base-pairing on the 5' end relative to the other strand.[72][73][74] The position of the stem-loop may also influence strand choice.[75] The other strand, called the passenger strand due to its lower levels in the steady state, is denoted with an asterisk (*) and is normally degraded. In some cases, both strands of the duplex are viable and become functional miRNA that target different mRNA populations.[76]
Members of the Argonaute (Ago) protein family are central to RISC function. Argonautes are needed for miRNA-induced silencing and contain two conserved RNA binding domains: a PAZ domain that can bind the single stranded 3' end of the mature miRNA and a PIWI domain that structurally resembles ribonuclease-H and functions to interact with the 5' end of the guide strand. They bind the mature miRNA and orient it for interaction with a target mRNA. Some argonautes, for example human Ago2, cleave target transcripts directly; argonautes may also recruit additional proteins to achieve translational repression.[77] The human genome encodes eight argonaute proteins divided by sequence similarities into two families: AGO (with four members present in all mammalian cells and called E1F2C/hAgo in humans), and PIWI (found in the germ line and hematopoietic stem cells).[71][77]
Additional RISC components include TRBP [human immunodeficiency virus (HIV) transactivating response RNA (TAR) binding protein],[78] PACT (protein activator of the interferon-induced protein kinase), the SMN complex, fragile X mental retardation protein (FMRP), Tudor staphylococcal nuclease-domain-containing protein (Tudor-SN), the putative DNA helicase MOV10, and the RNA recognition motif containing protein TNRC6B.[62][79][80]
Mode of silencing and regulatory loops
Gene silencing may occur either via mRNA degradation or preventing mRNA from being translated. For example, miR16 contains a sequence complementary to the AU-rich element found in the 3'UTR of many unstable mRNAs, such as TNF alpha or GM-CSF.[81] It has been demonstrated that given complete complementarity between the miRNA and target mRNA sequence, Ago2 can cleave the mRNA and lead to direct mRNA degradation. In the absence of complementarity, silencing is achieved by preventing translation.[40] The relation of miRNA and its target mRNA(s) can be based on the simple negative regulation of a target mRNA, but it seems that a common scenario is the use of a "coherent feed-forward loop", "mutual negative feedback loop" (also termed double negative loop) and "positive feedback/feed-forward loop". Some miRNAs work as buffers of random gene expression changes arising due to stochastic events in transcription, translation and protein stability. Such regulation is typically achieved by the virtue of negative feedback loops or incoherent feed-forward loop uncoupling protein output from mRNA transcription.
Turnover
Turnover of mature miRNA is needed for rapid changes in miRNA expression profiles. During miRNA maturation in the cytoplasm, uptake by the Argonaute protein is thought to stabilize the guide strand, while the opposite (* or "passenger") strand is preferentially destroyed. In what has been called a "Use it or lose it" strategy, Argonaute may preferentially retain miRNAs with many targets over miRNAs with few or no targets, leading to degradation of the non-targeting molecules.[82]
Decay of mature miRNAs in Caenorhabditis elegans is mediated by the 5´-to-3´ exoribonuclease XRN2, also known as Rat1p.[83] In plants, SDN (small RNA degrading nuclease) family members degrade miRNAs in the opposite (3'-to-5') direction. Similar enzymes are encoded in animal genomes, but their roles have not been described.[82]
Several miRNA modifications affect miRNA stability. As indicated by work in the model organism Arabidopsis thaliana (thale cress), mature plant miRNAs appear to be stabilized by the addition of methyl moieties at the 3' end. The 2'-O-conjugated methyl groups block the addition of uracil (U) residues by uridyltransferase enzymes, a modification that may be associated with miRNA degradation. However, uridylation may also protect some miRNAs; the consequences of this modification are incompletely understood. Uridylation of some animal miRNAs has been reported. Both plant and animal miRNAs may be altered by addition of adenine (A) residues to the 3' end of the miRNA. An extra A added to the end of mammalian miR-122, a liver-enriched miRNA important in hepatitis C, stabilizes the molecule and plant miRNAs ending with an adenine residue have slower decay rates.[82]
Cellular functions

Interaction of microRNA with protein translation process. Several translation repression mechanisms are shown: M1) on the initiation process, preventing assembling of the initiation complex or recruiting the 40S ribosomal subunit; M2) on the ribosome assembly; M3) on the translation process; M7, M8) on the degradation of mRNA.[84] 40S and 60S are light and heavy components of the ribosome, 80S is the assembled ribosome bound to mRNA, eIF4F is a translation initiation factor, PABC1 is the Poly-A binding protein, and "cap" is the mRNA cap structure needed for mRNA circularization (which can be the normal m7G-cap or modified A-cap). The initiation of mRNA can proceed in a cap-independent manner, through recruiting 40S to IRES (Internal Ribosome Entry Site) located in 5'UTR region. The actual work of RNA silencing is performed by RISC in which the main catalytic subunit is one of the Argonaute proteins (AGO), and miRNA serves as a template for recognizing specific mRNA sequences.
The function of miRNAs appears to be in gene regulation. For that purpose, a miRNA is complementary to a part of one or more messenger RNAs (mRNAs). Animal miRNAs are usually complementary to a site in the 3' UTR whereas plant miRNAs are usually complementary to coding regions of mRNAs.[85] Perfect or near perfect base pairing with the target RNA promotes cleavage of the RNA.[86] This is the primary mode of plant miRNAs.[87] In animals the match-ups are imperfect.
For partially complementary microRNAs to recognise their targets, nucleotides 2–7 of the miRNA (its 'seed region'[10][35]) must be perfectly complementary.[88] Animal miRNAs inhibit protein translation of the target mRNA[89] (this is present but less common in plants).[87] Partially complementary microRNAs can also speed up deadenylation, causing mRNAs to be degraded sooner.[90] While degradation of miRNA-targeted mRNA is well documented, whether or not translational repression is accomplished through mRNA degradation, translational inhibition, or a combination of the two is hotly debated. Recent work on miR-430 in zebrafish, as well as on bantam-miRNA and miR-9 in Drosophila cultured cells, shows that translational repression is caused by the disruption of translation initiation, independent of mRNA deadenylation.[91][92]
miRNAs occasionally also cause histone modification and DNA methylation of promoter sites, which affects the expression of target genes.[93][94]
Nine mechanisms of miRNA action are described and assembled in a unified mathematical model:[84]
- Cap-40S initiation inhibition;
- 60S Ribosomal unit joining inhibition;
- Elongation inhibition;
- Ribosome drop-off (premature termination);
- Co-translational nascent protein degradation;
- Sequestration in P-bodies;
- mRNA decay (destabilisation);
- mRNA cleavage;
- Transcriptional inhibition through microRNA-mediated chromatin reorganization followed by gene silencing.
It is often impossible to discern these mechanisms using experimental data about stationary reaction rates. Nevertheless, they are differentiated in dynamics and have different kinetic signatures.[84]
Unlike plant microRNAs, the animal microRNAs target diverse genes.[35] However, genes involved in functions common to all cells, such as gene expression, have relatively fewer microRNA target sites and seem to be under selection to avoid targeting by microRNAs.[95]
dsRNA can also activate gene expression, a mechanism that has been termed "small RNA-induced gene activation" or RNAa. dsRNAs targeting gene promoters can induce potent transcriptional activation of associated genes. This was demonstrated in human cells using synthetic dsRNAs termed small activating RNAs (saRNAs),[96] but has also been demonstrated for endogenous microRNA.[97]
Interactions between microRNAs and complementary sequences on genes and even pseudogenes that share sequence homology are thought to be a back channel of communication regulating expression levels between paralogous genes. Given the name "competing endogenous RNAs" (ceRNAs), these microRNAs bind to "microRNA response elements" on genes and pseudogenes and may provide another explanation for the persistence of non-coding DNA.[98]
Evolution
miRNAs are well conserved in both plants and animals, and are thought to be a vital and evolutionarily ancient component of gene regulation.[99][100][101][102][103] While core components of the microRNA pathway are conserved between plants and animals, miRNA repertoires in the two kingdoms appear to have emerged independently with different primary modes of action.[104][105]
microRNAs are useful phylogenetic markers because of their apparently low rate of evolution.[106] microRNAs' origin as a regulatory mechanism developed from previous RNAi machinery that was initially used as a defense against exogenous genetic material such as viruses.[107] Their origin may have permitted the development of morphological innovation, and by making gene expression more specific and 'fine-tunable', permitted the genesis of complex organs[108] and perhaps, ultimately, complex life.[103] Rapid bursts of morphological innovation are generally associated with a high rate of microRNA accumulation.[106][108]
New microRNAs are created in multiple ways. Novel microRNAs can originate from the random formation of hairpins in "non-coding" sections of DNA (i.e. introns or intergene regions), but also by the duplication and modification of existing microRNAs.[109] microRNAs can also form from inverted duplications of protein-coding sequences, which allows for the creation of a foldback hairpin structure.[110] The rate of evolution (i.e. nucleotide substitution) in recently originated microRNAs is comparable to that elsewhere in the non-coding DNA, implying evolution by neutral drift; however, older microRNAs have a much lower rate of change (often less than one substitution per hundred million years),[103] suggesting that once a microRNA gains a function, it undergoes purifying selection.[109] Individual regions within an miRNA gene face different evolutionary pressures, where regions that are vital for processing and function have higher levels of conservation.[111] At this point, a microRNA is rarely lost from an animal's genome,[103] although newer microRNAs (thus presumably non-functional) are frequently lost.[109] In Arabidopsis thaliana, the net flux of miRNA genes has been predicted to be between 1.2 and 3.3 genes per million years.[112] This makes them a valuable phylogenetic marker, and they are being looked upon as a possible solution to outstanding phylogenetic problems such as the relationships of arthropods.[113] On the other hand, in multiple cases microRNAs correlate poorly with phylogeny, and it is possible that their phylogenetic concordance largely reflects a limited sampling of microRNAs.[114]
microRNAs feature in the genomes of most eukaryotic organisms, from the brown algae[115] to the animals. However, the difference in how these microRNAs function and the way they are processed suggests that microRNAs arose independently in plants and animals.[116]
Focusing on the animals, the genome of Mnemiopsis leidyi[117] appears to lack recognizable microRNAs, as well as the nuclear proteins Drosha and Pasha, which are critical to canonical microRNA biogenesis. It is the only animal thus far reported to be missing Drosha. MicroRNAs play a vital role in the regulation of gene expression in all non-ctenophore animals investigated thus far except for Trichoplax adhaerens, the only known member of the phylum Placozoa.[118]
Across all species, in excess of 5000 different miRNAs had been identified by March 2010.[119] Whilst short RNA sequences (50 – hundreds of base pairs) of a broadly comparable function occur in bacteria, bacteria lack true microRNAs.[120]
Experimental detection and manipulation
While researchers focused on miRNA expression in physiological and pathological processes, various technical variables related to microRNA isolation emerged. The stability of stored miRNA samples has been questioned.[68] microRNAs degrade much more easily than mRNAs, partly due to their length, but also because of ubiquitously present RNases. This makes it necessary to cool samples on ice and use RNase-free equipment.[121]
microRNA expression can be quantified in a two-step polymerase chain reaction process of modified RT-PCR followed by quantitative PCR. Variations of this method achieve absolute or relative quantification.[122] miRNAs can also be hybridized to microarrays, slides or chips with probes to hundreds or thousands of miRNA targets, so that relative levels of miRNAs can be determined in different samples.[123] microRNAs can be both discovered and profiled by high-throughput sequencing methods (microRNA sequencing).[124] The activity of an miRNA can be experimentally inhibited using a locked nucleic acid (LNA) oligo, a Morpholino oligo[125][126] or a 2'-O-methyl RNA oligo.[127] A specific miRNA can be silenced by a complementary antagomir. microRNA maturation can be inhibited at several points by steric-blocking oligos.[128][129] The miRNA target site of an mRNA transcript can also be blocked by a steric-blocking oligo.[130] For the "in situ" detection of miRNA, LNA[131] or Morpholino[132] probes can be used. The locked conformation of LNA results in enhanced hybridization properties and increases sensitivity and selectivity, making it ideal for detection of short miRNA.[133]
High-throughput quantification of miRNAs is error prone, for the larger variance (compared to mRNAs) that comes with methodological problems. mRNA-expression is therefore often analyzed to check for miRNA-effects in their levels (e.g. in[134][135]). Databases can be used to pair mRNA- and miRNA-data that predict miRNA-targets based on their base sequence.[136][137] While this is usually done after miRNAs of interest have been detected (e. g. because of high expression levels), ideas for analysis tools that integrate mRNA- and miRNA-expression information have been proposed.[138][139]
Disease
Just as miRNA is involved in the normal functioning of eukaryotic cells, so has dysregulation of miRNA been associated with disease. A manually curated, publicly available database, miR2Disease, documents known relationships between miRNA dysregulation and human disease.[140]
Inherited diseases
A mutation in the seed region of miR-96, causes hereditary progressive hearing loss.[141]
A mutation in the seed region of miR-184, causes hereditary keratoconus with anterior polar cataract.[142]
Deletion of the miR-17~92 cluster, causes skeletal and growth defects.[143]
Cancer
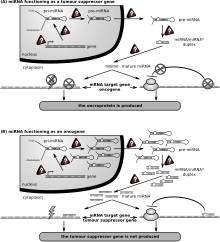
Role of miRNA in a cancer cell
The first human disease known to be associated with miRNA deregulation was chronic lymphocytic leukemia. Many other miRNAs also have links with cancer and accordingly are sometimes referred to as "oncomirs". In malignant B cells miRNAs participate in pathways fundamental to B cell development like B-cell receptor (BCR) signalling, B-cell migration/adhesion, cell-cell interactions in immune niches and the production and class-switching of immunoglobulins. MiRNAs influence B cell maturation, generation of pre-, marginal zone, follicular, B1, plasma and memory B cells.
Another role for miRNA in cancers is to use their expression level for prognosis. In NSCLC samples, low miR-324a levels may serve as an indicator of poor survival.[144] Either high miR-185 or low miR-133b levels may correlate with metastasis and poor survival in colorectal cancer.[145]
Furthermore, specific miRNAs may be associated with certain histological subtypes of colorectal cancer. For instance, expression levels of miR-205 and miR-373 have been shown to be increased in mucinous colorectal cancers and mucin-producing Ulcerative Colitis-associated colon cancers, but not in sporadic colonic adenocarcinoma that lack mucinous components.[146] In-vitro studies suggested that miR-205 and miR-373 may functionally induce different features of mucinous-associated neoplastic progression in intestinal epithelial cells.[146]
Hepatocellular carcinoma cell proliferation may arise from miR-21 interaction with MAP2K3, a tumor repressor gene.[147] Optimal treatment for cancer involves accurately identifying patients for risk-stratified therapy. Those with a rapid response to initial treatment may benefit from truncated treatment regimens, showing the value of accurate disease response measures. Cell-free miRNA are highly stable in blood, are overexpressed in cancer and are quantifiable within the diagnostic laboratory. In classical Hodgkin lymphoma, plasma miR-21, miR-494, and miR-1973 are promising disease response biomarkers.[148] Circulating miRNAs have the potential to assist clinical decision making and aid interpretation of positron emission tomography combined with computerized tomography. They can be performed at each consultation to assess disease response and detect relapse.
MicroRNAs have the potential to be used as tools or targets for treatment of different cancers.[149] The specific microRNA, miR-506 has been found to work as a tumor antagonist in several studies. A significant number of cervical cancer samples were found to have downregulated expression of miR-506. Additionally, miR-506 works to promote apoptosis of cervical cancer cells, through its direct target hedgehog pathway transcription factor, Gli3.[150][151]
DNA repair and cancer
DNA damage is considered to be the primary underlying cause of cancer.[152] If DNA repair is deficient, damage can accumulate. Such damage can cause mutational errors during DNA replication due to error-prone translesion synthesis. Accumulated damage can also cause epigenetic alterations due to errors during DNA repair.[153][154] Such mutations and epigenetic alterations can give rise to cancer (see malignant neoplasms).
Germ line mutations in DNA repair genes cause only 2–5% of colon cancer cases.[155] However, altered expression of microRNAs, causing DNA repair deficiencies, are frequently associated with cancers and may be an important causal factor.
Among 68 sporadic colon cancers with reduced expression of the DNA mismatch repair protein MLH1, most were found to be deficient due to epigenetic methylation of the CpG island of the MLH1 gene.[156] However, up to 15% of MLH1-deficiencies in sporadic colon cancers appeared to be due to over-expression of the microRNA miR-155, which represses MLH1 expression.[157]
In 29–66%[158][159] of glioblastomas, DNA repair is deficient due to epigenetic methylation of the MGMT gene, which reduces protein expression of MGMT. However, for 28% of glioblastomas, the MGMT protein is deficient, but the MGMT promoter is not methylated.[158] In glioblastomas without methylated MGMT promoters, the level of microRNA miR-181d is inversely correlated with protein expression of MGMT and the direct target of miR-181d is the MGMT mRNA 3'UTR (the three prime untranslated region of MGMT mRNA).[158] Thus, in 28% of glioblastomas, increased expression of miR-181d and reduced expression of DNA repair enzyme MGMT may be a causal factor.
HMGA proteins (HMGA1a, HMGA1b and HMGA2) are implicated in cancer, and expression of these proteins is regulated by microRNAs. HMGA expression is almost undetectable in differentiated adult tissues, but is elevated in many cancers. HMGA proteins are polypeptides of ~100 amino acid residues characterized by a modular sequence organization. These proteins have three highly positively charged regions, termed AT hooks, that bind the minor groove of AT-rich DNA stretches in specific regions of DNA. Human neoplasias, including thyroid, prostatic, cervical, colorectal, pancreatic and ovarian carcinomas, show a strong increase of HMGA1a and HMGA1b proteins.[160] Transgenic mice with HMGA1 targeted to lymphoid cells develop aggressive lymphoma, showing that high HMGA1 expression is associated with cancers and that HMGA1 can act as an oncogene.[161]
HMGA2 protein specifically targets the promoter of ERCC1, thus reducing expression of this DNA repair gene.[162] ERCC1 protein expression was deficient in 100% of 47 evaluated colon cancers (though the extent to which HGMA2 was involved is not known).[163]
Heart disease
The global role of miRNA function in the heart has been addressed by conditionally inhibiting miRNA maturation in the murine heart. This revealed that miRNAs play an essential role during its development.[164][165] miRNA expression profiling studies demonstrate that expression levels of specific miRNAs change in diseased human hearts, pointing to their involvement in cardiomyopathies.[166][167][168] Furthermore, animal studies on specific miRNAs identified distinct roles for miRNAs both during heart development and under pathological conditions, including the regulation of key factors important for cardiogenesis, the hypertrophic growth response and cardiac conductance.[165][169][170][171][172][173] Another role for miRNA in cardiovascular diseases is to use their expression levels for diagnosis, prognosis or risk stratification.[174] miRNA's in animal models have also been linked to cholesterol metabolism and regulation.[175]
miRNA-712
Murine microRNA-712 is a potential biomarker (i.e. predictor) for atherosclerosis, a cardiovascular disease of the arterial wall associated with lipid retention and inflammation.[176] Non-laminar blood flow also correlates with development of atherosclerosis as mechanosenors of endothelial cells respond to the shear force of disturbed flow (d-flow).[177] A number of pro-atherogenic genes including matrix metalloproteinases (MMPs) are upregulated by d-flow ,[177] mediating pro-inflammatory and pro-angiogenic signals. These findings were observed in ligated carotid arteries of mice to mimic the effects of d-flow. Within 24 hours, pre-existing immature miR-712 formed mature miR-712 suggesting that miR-712 is flow-sensitive.[177] Coinciding with these results, miR-712 is also upregulated in endothelial cells exposed to naturally occurring d-flow in the greater curvature of the aortic arch.[177]
Gene origin
Pre-mRNA sequence of miR-712 is generated from the murine ribosomal RN45s gene at the internal transcribed spacer region 2 (ITS2).[177] XRN1 is an exonuclease that degrades the ITS2 region during processing of RN45s.[177] Reduction of XRN1 under d-flow conditions therefore leads to the accumulation of miR-712.[177]
Mechanism
MiR-712 targets tissue inhibitor of metalloproteinases 3 (TIMP3).[177] TIMPs normally regulate activity of matrix metalloproteinases (MMPs) which degrade the extracellular matrix (ECM). Arterial ECM is mainly composed of collagen and elastin fibers, providing the structural support and recoil properties of arteries.[178] These fibers play a critical role in regulation of vascular inflammation and permeability, which are important in the development of atherosclerosis.[179] Expressed by endothelial cells, TIMP3 is the only ECM-bound TIMP.[178] A decrease in TIMP3 expression results in an increase of ECM degradation in the presence of d-flow. Consistent with these findings, inhibition of pre-miR712 increases expression of TIMP3 in cells, even when exposed to turbulent flow.[177]
TIMP3 also decreases the expression of TNFα (a pro-inflammatory regulator) during turbulent flow.[177] Activity of TNFα in turbulent flow was measured by the expression of TNFα-converting enzyme (TACE) in blood. TNFα decreased if miR-712 was inhibited or TIMP3 overexpressed,[177] suggesting that miR-712 and TIMP3 regulate TACE activity in turbulent flow conditions.
Anti-miR-712 effectively suppresses d-flow-induced miR-712 expression and increases TIMP3 expression.[177] Anti-miR-712 also inhibits vascular hyperpermeability, thereby significantly reducing atherosclerosis lesion development and immune cell infiltration.[177]
Human homolog microRNA-205
The human homolog of miR-712 was found on the RN45s homolog gene, which maintains similar miRNAs to mice.[177] MiR-205 of humans sshare similar sequences with miR-712 of mice and is conserved across most vertebrates.[177] MiR-205 and miR-712 also share more than 50% of the cell signaling targets, including TIMP3.[177]
When tested, d-flow decreased the expression of XRN1 in humans as it did in mice endothelial cells, indicating a potentially common role of XRN1 in humans.[177]
Kidney disease
Targeted deletion of Dicer in the FoxD1-derived renal progenitor cells in a murine model resulted in a complex renal phenotype including expansion of nephron progenitors, fewer renin cells, smooth muscle arterioles, progressive mesangial loss and glomerular aneurysms.[180] High throughput whole transcriptome profiling of the FoxD1-Dicer knockout mouse model revealed ectopic upregulation of pro-apoptotic gene, Bcl2L11 (Bim) and dysregulation of the p53 pathway with increase in p53 effector genes including Bax, Trp53inp1, Jun, Cdkn1a, Mmp2, and Arid3a. p53 protein levels remained unchanged, suggesting that FoxD1 stromal miRNAs directly repress p53-effector genes. Using a lineage tracing approach followed by Fluorescent-activated cell sorting, miRNA profiling of the FoxD1-derived cells not only comprehensively defined the transcriptional landscape of miRNAs that are critical for vascular development, but also identified key miRNAs that are likely to modulate the renal phenotype in its absence. These miRNAs include miRs‐10a, 18a, 19b, 24, 30c, 92a, 106a, 130a, 152, 181a, 214, 222, 302a, 370, and 381 that regulate Bcl2L11 (Bim) and miRs‐15b, 18a, 21, 30c, 92a, 106a, 125b‐5p, 145, 214, 222, 296‐5p and 302a that regulate p53-effector genes. Consistent with the profiling results, ectopic apoptosis was observed in the cellular derivatives of the FoxD1 derived progenitor lineage and reiterates the importance of renal stromal miRNAs in cellular homeostasis.[180]
Nervous system
miRNAs appear to regulate the development and function of the nervous system.[181] Neural miRNAs are involved at various stages of synaptic development, including dendritogenesis (involving miR-132, miR-134 and miR-124), synapse formation[182] and synapse maturation (where miR-134 and miR-138 are thought to be involved).[183] Some studies find altered miRNA expression in schizophrenia, as well as bipolar disorder and major depression and anxiety disorders.[184][185][186]
Alcoholism
The vital role of miRNAs in gene expression is significant to addiction, specifically alcoholism.[187] Chronic alcohol abuse results in persistent changes in brain function mediated in part by alterations in gene expression.[187] miRNA global regulation of many downstream genes deems significant regarding the reorganization or synaptic connections or long term neuroadaptions involving the behavioral change from alcohol consumption to withdrawal and/or dependence.[188] Up to 35 different miRNAs have been found to be altered in the alcoholic post-mortem brain, all of which target genes that include the regulation of the cell cycle, apoptosis, cell adhesion, nervous system development and cell signaling.[187] Altered miRNA levels were found in the medial prefrontal cortex of alcohol-dependent mice, suggesting the role of miRNA in orchestrating translational imbalances and the creation of differentially expressed proteins within an area of the brain where complex cognitive behavior and decision making likely originate.[189]
miRNAs can be either upregulated or downregulated in response to chronic alcohol use. miR-206 expression increased in the prefrontal cortex of alcohol-dependent rats, targeting the transcription factor brain-derived neurotrophic factor (BDNF) and ultimately reducing its expression. BDNF plays a critical role in the formation and maturation of new neurons and synapses, suggesting a possible implication in synapse growth/synaptic plasticity in alcohol abusers.[190] miR-155, important in regulating alcohol-induced neuroinflammation responses, was found to be upregulated, suggesting the role of microglia and inflammatory cytokines in alcohol pathophysiology.[191] Downregulation of miR-382 was found in the nucleus accumbens, a structure in the basal forebrain significant in regulating feelings of reward that power motivational habits. miR-382 is the target for the dopamine receptor D1 (DRD1), and its overexpression results in the upregulation of DRD1 and delta fosB, a transcription factor that activates a series of transcription events in the nucleus accumbens that ultimately result in addictive behaviors.[192] Alternatively, overexpressing miR-382 resulted in attenuated drinking and the inhibition of DRD1 and delta fosB upregulation in rat models of alcoholism, demonstrating the possibility of using miRNA-targeted pharmaceuticals in treatments.[192]
Obesity
miRNAs play crucial roles in the regulation of stem cell progenitors differentiating into adipocytes.[193] Studies to determine what role pluripotent stem cells play in adipogenesis, were examined in the immortalized human bone marrow-derived stromal cell line hMSC-Tert20.[194] Decreased expression of miR-155, miR-221, and miR-222, have been found during the adipogenic programming of both immortalized and primary hMSCs, suggesting that they act as negative regulators of differentiation. Conversely, ectopic expression of the miRNAs 155,221, and 222 significantly inhibited adipogenesis and repressed induction of the master regulators PPARγ and CCAAT/enhancer-binding protein alpha (CEBPA).[195] This paves the way for possible genetic obesity treatments.
Another class of miRNAs that regulate insulin resistance, obesity, and diabetes, is the let-7 family. Let-7 accumulates in human tissues during the course of aging.[196] When let-7 was ectopically overexpressed to mimic accelerated aging, mice became insulin-resistant, and thus more prone to high fat diet-induced obesity and diabetes.[197] In contrast when let-7 was inhibited by injections of let-7-specific antagomirs, mice become more insulin-sensitive and remarkably resistant to high fat diet-induced obesity and diabetes. Not only could let-7 inhibition prevent obesity and diabetes, it could also reverse and cure the condition.[198] These experimental findings suggest that let-7 inhibition could represent a new therapy for obesity and type 2 diabetes.
Hemostasis
miRNAs also play crucial roles in the regulation of complex enzymatic cascades including the hemostatic blood coagulation system.[199] Large scale studies of functional miRNA targeting have recently uncovered rationale therapeutic targets in the hemostatic system.[200]
Non-coding RNAs
When the human genome project mapped its first chromosome in 1999, it was predicted the genome would contain over 100,000 protein coding genes. However, only around 20,000 were eventually identified.[201] Since then, the advent of bioinformatics approaches combined with genome tiling studies examining the transcriptome,[202] systematic sequencing of full length cDNA libraries[203] and experimental validation[204] (including the creation of miRNA derived antisense oligonucleotides called antagomirs) have revealed that many transcripts are non protein-coding RNA, including several snoRNAs and miRNAs.[205]
Viruses
Viral microRNAs play an important role in the regulation of gene expression of viral and/or host genes to benefit the virus. Hence, miRNAs play a key role in host–virus interactions and pathogenesis of viral diseases.[206][207] The expression of transcription activators by human herpesvirus-6 DNA is believed to be regulated by viral miRNA.[208]
Target prediction
miRNAs can bind to target messenger RNA (mRNA) transcripts of protein-coding genes and negatively control their translation or cause mRNA degradation. It is of key importance to identify the miRNA targets accurately.[209] A comparison of the predictive performance of eighteen in silico algorithms is available.[210] Large scale studies of functional miRNA targeting suggest that many functional miRNAs can be missed by target prediction algorithms.[200]
See also
- Gene expression
- List of miRNA gene prediction tools
- List of miRNA target prediction tools
- RNA interference
- Small interfering RNA
- Small nucleolar RNA-derived microRNA
- Anti-miRNA oligonucleotides
- MicroDNA
- miR-324-5p
References
^ Ambros V (September 2004). "The functions of animal microRNAs". Nature. 431 (7006): 350–5. Bibcode:2004Natur.431..350A. doi:10.1038/nature02871. PMID 15372042..mw-parser-output cite.citationfont-style:inherit.mw-parser-output qquotes:"""""""'""'".mw-parser-output code.cs1-codecolor:inherit;background:inherit;border:inherit;padding:inherit.mw-parser-output .cs1-lock-free abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-lock-subscription abackground:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registrationcolor:#555.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration spanborder-bottom:1px dotted;cursor:help.mw-parser-output .cs1-hidden-errordisplay:none;font-size:100%.mw-parser-output .cs1-visible-errorfont-size:100%.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-formatfont-size:95%.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-leftpadding-left:0.2em.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-rightpadding-right:0.2em
^ abc Bartel DP (January 2004). "MicroRNAs: genomics, biogenesis, mechanism, and function". Cell. 116 (2): 281–97. doi:10.1016/S0092-8674(04)00045-5. PMID 14744438.
^ abc Bartel DP (March 2018). "Metazoan MicroRNAs". Cell. 173 (1): 20–51. doi:10.1016/j.cell.2018.03.006. PMC 6091663. PMID 29570994.
^ abcd Bartel DP (January 2009). "MicroRNAs: target recognition and regulatory functions". Cell. 136 (2): 215–33. doi:10.1016/j.cell.2009.01.002. PMC 3794896. PMID 19167326.
^ Fabian MR, Sonenberg N, Filipowicz W (2010). "Regulation of mRNA translation and stability by microRNAs". Annual Review of Biochemistry. 79: 351–79. doi:10.1146/annurev-biochem-060308-103103. PMID 20533884.
^ Homo sapiens miRNAs in the miRBase at Manchester University
^ ab Fromm B, Billipp T, Peck LE, Johansen M, Tarver JE, King BL, et al. (2015). "A Uniform System for the Annotation of Vertebrate microRNA Genes and the Evolution of the Human microRNAome". Annual Review of Genetics. 49: 213–42. doi:10.1146/annurev-genet-120213-092023. PMC 4743252. PMID 26473382.
^ Lim LP, Lau NC, Weinstein EG, Abdelhakim A, Yekta S, Rhoades MW, Burge CB, Bartel DP (April 2003). "The microRNAs of Caenorhabditis elegans". Genes & Development. 17 (8): 991–1008. doi:10.1101/gad.1074403. PMC 196042. PMID 12672692.
^ ab Lagos-Quintana M, Rauhut R, Yalcin A, Meyer J, Lendeckel W, Tuschl T (April 2002). "Identification of tissue-specific microRNAs from mouse". Current Biology. 12 (9): 735–9. doi:10.1016/S0960-9822(02)00809-6. PMID 12007417.
^ abc Lewis BP, Burge CB, Bartel DP (January 2005). "Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets". Cell. 120 (1): 15–20. doi:10.1016/j.cell.2004.12.035. PMID 15652477.
^ abc Friedman RC, Farh KK, Burge CB, Bartel DP (January 2009). "Most mammalian mRNAs are conserved targets of microRNAs". Genome Research. 19 (1): 92–105. doi:10.1101/gr.082701.108. PMC 2612969. PMID 18955434.
^ abc Lee RC, Feinbaum RL, Ambros V (December 1993). "The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14". Cell. 75 (5): 843–54. doi:10.1016/0092-8674(93)90529-Y. PMID 8252621.
^ ab Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR, Ruvkun G (February 2000). "The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans". Nature. 403 (6772): 901–6. Bibcode:2000Natur.403..901R. doi:10.1038/35002607. PMID 10706289.
^ ab Pasquinelli AE, Reinhart BJ, Slack F, Martindale MQ, Kuroda MI, Maller B, Hayward DC, Ball EE, Degnan B, Müller P, Spring J, Srinivasan A, Fishman M, Finnerty J, Corbo J, Levine M, Leahy P, Davidson E, Ruvkun G (November 2000). "Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA". Nature. 408 (6808): 86–9. doi:10.1038/35040556. PMID 11081512.
^ abc Lagos-Quintana M, Rauhut R, Lendeckel W, Tuschl T (October 2001). "Identification of novel genes coding for small expressed RNAs". Science. 294 (5543): 853–8. Bibcode:2001Sci...294..853L. doi:10.1126/science.1064921. PMID 11679670.
^ abc Lau NC, Lim LP, Weinstein EG, Bartel DP (October 2001). "An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans". Science. 294 (5543): 858–62. Bibcode:2001Sci...294..858L. doi:10.1126/science.1065062. PMID 11679671.
^ abc Lee RC, Ambros V (October 2001). "An extensive class of small RNAs in Caenorhabditis elegans". Science. 294 (5543): 862–4. Bibcode:2001Sci...294..862L. doi:10.1126/science.1065329. PMID 11679672.
^ Wienholds E, Kloosterman WP, Miska E, Alvarez-Saavedra E, Berezikov E, de Bruijn E, Horvitz HR, Kauppinen S, Plasterk RH (July 2005). "MicroRNA expression in zebrafish embryonic development". Science. 309 (5732): 310–1. Bibcode:2005Sci...309..310W. doi:10.1126/science.1114519. PMID 15919954.
^ ab Jones-Rhoades MW, Bartel DP, Bartel B (2006). "MicroRNAS and their regulatory roles in plants". Annual Review of Plant Biology. 57: 19–53. doi:10.1146/annurev.arplant.57.032905.105218. PMID 16669754.
^ Brennecke J, Hipfner DR, Stark A, Russell RB, Cohen SM (April 2003). "bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila". Cell. 113 (1): 25–36. doi:10.1016/S0092-8674(03)00231-9. PMID 12679032.
^ Zhang J, Qu P, Zhou C, Liu X, Ma X, Wang M, Wang Y, Su J, Liu J, Zhang Y (September 2017). "MicroRNA-125b is a key epigenetic regulatory factor that promotes nuclear transfer reprogramming". The Journal of Biological Chemistry. 292 (38): 15916–15926. doi:10.1074/jbc.M117.796771. PMC 5612121. PMID 28794155.
^ Cuellar TL, McManus MT (December 2005). "MicroRNAs and endocrine biology". The Journal of Endocrinology. 187 (3): 327–32. doi:10.1677/joe.1.06426. PMID 16423811.
^ Poy MN, Eliasson L, Krutzfeldt J, Kuwajima S, Ma X, Macdonald PE, Pfeffer S, Tuschl T, Rajewsky N, Rorsman P, Stoffel M (November 2004). "A pancreatic islet-specific microRNA regulates insulin secretion". Nature. 432 (7014): 226–30. Bibcode:2004Natur.432..226P. doi:10.1038/nature03076. PMID 15538371.
^ Chen CZ, Li L, Lodish HF, Bartel DP (January 2004). "MicroRNAs modulate hematopoietic lineage differentiation". Science. 303 (5654): 83–6. Bibcode:2004Sci...303...83C. doi:10.1126/science.1091903. hdl:1721.1/7483. PMID 14657504.
^ Wilfred BR, Wang WX, Nelson PT (July 2007). "Energizing miRNA research: a review of the role of miRNAs in lipid metabolism, with a prediction that miR-103/107 regulates human metabolic pathways". Molecular Genetics and Metabolism. 91 (3): 209–17. doi:10.1016/j.ymgme.2007.03.011. PMC 1978064. PMID 17521938.
^ Harfe BD, McManus MT, Mansfield JH, Hornstein E, Tabin CJ (August 2005). "The RNaseIII enzyme Dicer is required for morphogenesis but not patterning of the vertebrate limb". Proceedings of the National Academy of Sciences of the United States of America. 102 (31): 10898–903. Bibcode:2005PNAS..10210898H. doi:10.1073/pnas.0504834102. PMC 1182454. PMID 16040801.
^ Trang P, Weidhaas JB, Slack FJ (December 2008). "MicroRNAs as potential cancer therapeutics". Oncogene. 27 Suppl 2: S52–7. doi:10.1038/onc.2009.353. PMID 19956180.
^ Li C, Feng Y, Coukos G, Zhang L (December 2009). "Therapeutic microRNA strategies in human cancer". The AAPS Journal. 11 (4): 747–57. doi:10.1208/s12248-009-9145-9. PMC 2782079. PMID 19876744.
^ Fasanaro P, Greco S, Ivan M, Capogrossi MC, Martelli F (January 2010). "microRNA: emerging therapeutic targets in acute ischemic diseases". Pharmacology & Therapeutics. 125 (1): 92–104. doi:10.1016/j.pharmthera.2009.10.003. PMID 19896977.
^ Hydbring P, Badalian-Very G (August 2013). "Clinical applications of microRNAs". F1000Research. 2: 136. doi:10.12688/f1000research.2-136.v2. PMC 3917658. PMID 24627783.
^ Wightman B, Ha I, Ruvkun G (December 1993). "Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans". Cell. 75 (5): 855–62. doi:10.1016/0092-8674(93)90530-4. PMID 8252622.
^ Ambros V, Bartel B, Bartel DP, Burge CB, Carrington JC, Chen X, Dreyfuss G, Eddy SR, Griffiths-Jones S, Marshall M, Matzke M, Ruvkun G, Tuschl T (March 2003). "A uniform system for microRNA annotation". RNA. 9 (3): 277–9. doi:10.1261/rna.2183803. PMC 1370393. PMID 12592000.
^ Griffiths-Jones S, Grocock RJ, van Dongen S, Bateman A, Enright AJ (January 2006). "miRBase: microRNA sequences, targets and gene nomenclature". Nucleic Acids Research. 34 (Database issue): D140–4. doi:10.1093/nar/gkj112. PMC 1347474. PMID 16381832.
^ Wright MW, Bruford EA (January 2011). "Naming 'junk': human non-protein coding RNA (ncRNA) gene nomenclature". Human Genomics. 5 (2): 90–8. doi:10.1186/1479-7364-5-2-90. PMC 3051107. PMID 21296742.
^ abc Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB (December 2003). "Prediction of mammalian microRNA targets". Cell. 115 (7): 787–98. doi:10.1016/S0092-8674(03)01018-3. PMID 14697198.
^ Ellwanger DC, Büttner FA, Mewes HW, Stümpflen V (May 2011). "The sufficient minimal set of miRNA seed types". Bioinformatics. 27 (10): 1346–50. doi:10.1093/bioinformatics/btr149. PMC 3087955. PMID 21441577.
^ Rajewsky N (June 2006). "microRNA target predictions in animals". Nature Genetics. 38 Suppl (6s): S8–13. doi:10.1038/ng1798. PMID 16736023.
^ Krek A, Grün D, Poy MN, Wolf R, Rosenberg L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M, Rajewsky N (May 2005). "Combinatorial microRNA target predictions". Nature Genetics. 37 (5): 495–500. doi:10.1038/ng1536. PMID 15806104.
^ Thomson DW, Bracken CP, Goodall GJ (September 2011). "Experimental strategies for microRNA target identification". Nucleic Acids Research. 39 (16): 6845–53. doi:10.1093/nar/gkr330. PMC 3167600. PMID 21652644.
^ ab Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, Bartel DP, Linsley PS, Johnson JM (February 2005). "Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs". Nature. 433 (7027): 769–73. Bibcode:2005Natur.433..769L. doi:10.1038/nature03315. PMID 15685193.
^ Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N (September 2008). "Widespread changes in protein synthesis induced by microRNAs". Nature. 455 (7209): 58–63. Bibcode:2008Natur.455...58S. doi:10.1038/nature07228. PMID 18668040.
^ Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP (September 2008). "The impact of microRNAs on protein output". Nature. 455 (7209): 64–71. Bibcode:2008Natur.455...64B. doi:10.1038/nature07242. PMC 2745094. PMID 18668037.
^ ab Rodriguez A, Griffiths-Jones S, Ashurst JL, Bradley A (October 2004). "Identification of mammalian microRNA host genes and transcription units". Genome Research. 14 (10A): 1902–10. doi:10.1101/gr.2722704. PMC 524413. PMID 15364901.
^ abcd Cai X, Hagedorn CH, Cullen BR (December 2004). "Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs". RNA. 10 (12): 1957–66. doi:10.1261/rna.7135204. PMC 1370684. PMID 15525708.
^ Weber MJ (January 2005). "New human and mouse microRNA genes found by homology search". The FEBS Journal. 272 (1): 59–73. doi:10.1111/j.1432-1033.2004.04389.x. PMID 15634332.
^ Kim YK, Kim VN (February 2007). "Processing of intronic microRNAs". The EMBO Journal. 26 (3): 775–83. doi:10.1038/sj.emboj.7601512. PMC 1794378. PMID 17255951.
^ Baskerville S, Bartel DP (March 2005). "Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes". RNA. 11 (3): 241–7. doi:10.1261/rna.7240905. PMC 1370713. PMID 15701730.
^ abc Lee Y, Kim M, Han J, Yeom KH, Lee S, Baek SH, Kim VN (October 2004). "MicroRNA genes are transcribed by RNA polymerase II". The EMBO Journal. 23 (20): 4051–60. doi:10.1038/sj.emboj.7600385. PMC 524334. PMID 15372072.
^ Zhou X, Ruan J, Wang G, Zhang W (March 2007). "Characterization and identification of microRNA core promoters in four model species". PLoS Computational Biology. 3 (3): e37. Bibcode:2007PLSCB...3...37Z. doi:10.1371/journal.pcbi.0030037. PMC 1817659. PMID 17352530.
^ Faller M, Guo F (November 2008). "MicroRNA biogenesis: there's more than one way to skin a cat". Biochimica et Biophysica Acta. 1779 (11): 663–7. doi:10.1016/j.bbagrm.2008.08.005. PMC 2633599. PMID 18778799.
^ Lee Y, Ahn C, Han J, Choi H, Kim J, Yim J, Lee J, Provost P, Rådmark O, Kim S, Kim VN (September 2003). "The nuclear RNase III Drosha initiates microRNA processing". Nature. 425 (6956): 415–9. Bibcode:2003Natur.425..415L. doi:10.1038/nature01957. PMID 14508493.
^ Gregory RI, Chendrimada TP, Shiekhattar R (2006). MicroRNA biogenesis: isolation and characterization of the microprocessor complex. Methods in Molecular Biology. 342. pp. 33–47. doi:10.1385/1-59745-123-1:33. ISBN 978-1-59745-123-9. PMID 16957365.
^ Han J, Lee Y, Yeom KH, Kim YK, Jin H, Kim VN (December 2004). "The Drosha-DGCR8 complex in primary microRNA processing". Genes & Development. 18 (24): 3016–27. doi:10.1101/gad.1262504. PMC 535913. PMID 15574589.
^ Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK, Sohn SY, Cho Y, Zhang BT, Kim VN (June 2006). "Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex". Cell. 125 (5): 887–901. doi:10.1016/j.cell.2006.03.043. PMID 16751099.
^ Conrad T, Marsico A, Gehre M, Orom UA (October 2014). "Microprocessor activity controls differential miRNA biogenesis In Vivo". Cell Reports. 9 (2): 542–54. doi:10.1016/j.celrep.2014.09.007. PMID 25310978.
^ Auyeung VC, Ulitsky I, McGeary SE, Bartel DP (February 2013). "Beyond secondary structure: primary-sequence determinants license pri-miRNA hairpins for processing". Cell. 152 (4): 844–58. doi:10.1016/j.cell.2013.01.031. PMC 3707628. PMID 23415231.
^ Ali PS, Ghoshdastider U, Hoffmann J, Brutschy B, Filipek S (November 2012). "Recognition of the let-7g miRNA precursor by human Lin28B". FEBS Letters. 586 (22): 3986–90. doi:10.1016/j.febslet.2012.09.034. PMID 23063642.
^ Berezikov E, Chung WJ, Willis J, Cuppen E, Lai EC (October 2007). "Mammalian mirtron genes". Molecular Cell. 28 (2): 328–36. doi:10.1016/j.molcel.2007.09.028. PMC 2763384. PMID 17964270.
^ ab Kawahara Y, Megraw M, Kreider E, Iizasa H, Valente L, Hatzigeorgiou AG, Nishikura K (September 2008). "Frequency and fate of microRNA editing in human brain". Nucleic Acids Research. 36 (16): 5270–80. doi:10.1093/nar/gkn479. PMC 2532740. PMID 18684997.
^ Winter J, Jung S, Keller S, Gregory RI, Diederichs S (March 2009). "Many roads to maturity: microRNA biogenesis pathways and their regulation". Nature Cell Biology. 11 (3): 228–34. doi:10.1038/ncb0309-228. PMID 19255566.
^ Ohman M (October 2007). "A-to-I editing challenger or ally to the microRNA process". Biochimie. 89 (10): 1171–6. doi:10.1016/j.biochi.2007.06.002. PMID 17628290.
^ ab Murchison EP, Hannon GJ (June 2004). "miRNAs on the move: miRNA biogenesis and the RNAi machinery". Current Opinion in Cell Biology. 16 (3): 223–9. doi:10.1016/j.ceb.2004.04.003. PMID 15145345.
^ abc Lund E, Dahlberg JE (2006). "Substrate selectivity of exportin 5 and Dicer in the biogenesis of microRNAs". Cold Spring Harbor Symposia on Quantitative Biology. 71: 59–66. doi:10.1101/sqb.2006.71.050. PMID 17381281.
^ Park JE, Heo I, Tian Y, Simanshu DK, Chang H, Jee D, Patel DJ, Kim VN (July 2011). "Dicer recognizes the 5' end of RNA for efficient and accurate processing". Nature. 475 (7355): 201–5. doi:10.1038/nature10198. PMC 4693635. PMID 21753850.
^ Ji X (2008). The mechanism of RNase III action: how dicer dices. Current Topics in Microbiology and Immunology. 320. pp. 99–116. doi:10.1007/978-3-540-75157-1_5. ISBN 978-3-540-75156-4. PMID 18268841.
^ Mirihana Arachchilage G, Dassanayake AC, Basu S (February 2015). "A potassium ion-dependent RNA structural switch regulates human pre-miRNA 92b maturation". Chemistry & Biology. 22 (2): 262–72. doi:10.1016/j.chembiol.2014.12.013. PMID 25641166.
^ Sohel MH (2016). "Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges". Achievements in the Life Sciences. 10 (2): 175–186. doi:10.1016/j.als.2016.11.007.
^ ab Boeckel JN, Reis SM, Leistner D, Thomé CE, Zeiher AM, Fichtlscherer S, Keller T (April 2014). "From heart to toe: heart's contribution on peripheral microRNA levels". International Journal of Cardiology. 172 (3): 616–7. doi:10.1016/j.ijcard.2014.01.082. PMID 24508494.
^ Lelandais-Brière C, Sorin C, Declerck M, Benslimane A, Crespi M, Hartmann C (March 2010). "Small RNA diversity in plants and its impact in development". Current Genomics. 11 (1): 14–23. doi:10.2174/138920210790217918. PMC 2851111. PMID 20808519.
^ Rana TM (January 2007). "Illuminating the silence: understanding the structure and function of small RNAs". Nature Reviews. Molecular Cell Biology. 8 (1): 23–36. doi:10.1038/nrm2085. PMID 17183358.
^ ab Schwarz DS, Zamore PD (May 2002). "Why do miRNAs live in the miRNP?". Genes & Development. 16 (9): 1025–31. doi:10.1101/gad.992502. PMID 12000786.
^ Krol J, Sobczak K, Wilczynska U, Drath M, Jasinska A, Kaczynska D, Krzyzosiak WJ (October 2004). "Structural features of microRNA (miRNA) precursors and their relevance to miRNA biogenesis and small interfering RNA/short hairpin RNA design". The Journal of Biological Chemistry. 279 (40): 42230–9. doi:10.1074/jbc.M404931200. PMID 15292246.
^ Khvorova A, Reynolds A, Jayasena SD (October 2003). "Functional siRNAs and miRNAs exhibit strand bias". Cell. 115 (2): 209–16. doi:10.1016/S0092-8674(03)00801-8. PMID 14567918.
^ Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (October 2003). "Asymmetry in the assembly of the RNAi enzyme complex". Cell. 115 (2): 199–208. doi:10.1016/S0092-8674(03)00759-1. PMID 14567917.
^ Lin SL, Chang D, Ying SY (August 2005). "Asymmetry of intronic pre-miRNA structures in functional RISC assembly". Gene. 356: 32–8. doi:10.1016/j.gene.2005.04.036. PMC 1788082. PMID 16005165.
^ Okamura K, Chung WJ, Lai EC (September 2008). "The long and short of inverted repeat genes in animals: microRNAs, mirtrons and hairpin RNAs". Cell Cycle. 7 (18): 2840–5. doi:10.4161/cc.7.18.6734. PMC 2697033. PMID 18769156.
^ ab Pratt AJ, MacRae IJ (July 2009). "The RNA-induced silencing complex: a versatile gene-silencing machine". The Journal of Biological Chemistry. 284 (27): 17897–901. doi:10.1074/jbc.R900012200. PMC 2709356. PMID 19342379.
^ MacRae IJ, Ma E, Zhou M, Robinson CV, Doudna JA (January 2008). "In vitro reconstitution of the human RISC-loading complex". Proceedings of the National Academy of Sciences of the United States of America. 105 (2): 512–7. Bibcode:2008PNAS..105..512M. doi:10.1073/pnas.0710869105. PMC 2206567. PMID 18178619.
^ Mourelatos Z, Dostie J, Paushkin S, Sharma A, Charroux B, Abel L, Rappsilber J, Mann M, Dreyfuss G (March 2002). "miRNPs: a novel class of ribonucleoproteins containing numerous microRNAs". Genes & Development. 16 (6): 720–8. doi:10.1101/gad.974702. PMC 155365. PMID 11914277.
^ Meister G, Landthaler M, Peters L, Chen PY, Urlaub H, Lührmann R, Tuschl T (December 2005). "Identification of novel argonaute-associated proteins". Current Biology. 15 (23): 2149–55. doi:10.1016/j.cub.2005.10.048. PMID 16289642.
^ Jing Q, Huang S, Guth S, Zarubin T, Motoyama A, Chen J, Di Padova F, Lin SC, Gram H, Han J (March 2005). "Involvement of microRNA in AU-rich element-mediated mRNA instability". Cell. 120 (5): 623–34. doi:10.1016/j.cell.2004.12.038. PMID 15766526.
^ abc Kai ZS, Pasquinelli AE (January 2010). "MicroRNA assassins: factors that regulate the disappearance of miRNAs". Nature Structural & Molecular Biology. 17 (1): 5–10. doi:10.1038/nsmb.1762. PMID 20051982.
^ Chatterjee S, Grosshans H (September 2009). "Active turnover modulates mature microRNA activity in Caenorhabditis elegans". Nature. 461 (7263): 546–9. Bibcode:2009Natur.461..546C. doi:10.1038/nature08349. PMID 19734881.
^ abc Morozova N, Zinovyev A, Nonne N, Pritchard LL, Gorban AN, Harel-Bellan A (September 2012). "Kinetic signatures of microRNA modes of action". RNA. 18 (9): 1635–55. doi:10.1261/rna.032284.112. PMC 3425779. PMID 22850425.
^ Wang XJ, Reyes JL, Chua NH, Gaasterland T (2004). "Prediction and identification of Arabidopsis thaliana microRNAs and their mRNA targets". Genome Biology. 5 (9): R65. doi:10.1186/gb-2004-5-9-r65. PMC 522872. PMID 15345049.
^ Kawasaki H, Taira K (2004). "MicroRNA-196 inhibits HOXB8 expression in myeloid differentiation of HL60 cells". Nucleic Acids Symposium Series. 48 (48): 211–2. doi:10.1093/nass/48.1.211. PMID 17150553.
^ ab Moxon S, Jing R, Szittya G, Schwach F, Rusholme Pilcher RL, Moulton V, Dalmay T (October 2008). "Deep sequencing of tomato short RNAs identifies microRNAs targeting genes involved in fruit ripening". Genome Research. 18 (10): 1602–9. doi:10.1101/gr.080127.108. PMC 2556272. PMID 18653800.
^ Mazière P, Enright AJ (June 2007). "Prediction of microRNA targets". Drug Discovery Today. 12 (11–12): 452–8. doi:10.1016/j.drudis.2007.04.002. PMID 17532529.
^ Williams AE (February 2008). "Functional aspects of animal microRNAs". Cellular and Molecular Life Sciences. 65 (4): 545–62. doi:10.1007/s00018-007-7355-9. PMID 17965831.
^ Eulalio A, Huntzinger E, Nishihara T, Rehwinkel J, Fauser M, Izaurralde E (January 2009). "Deadenylation is a widespread effect of miRNA regulation". RNA. 15 (1): 21–32. doi:10.1261/rna.1399509. PMC 2612776. PMID 19029310.
^ Bazzini AA, Lee MT, Giraldez AJ (April 2012). "Ribosome profiling shows that miR-430 reduces translation before causing mRNA decay in zebrafish". Science. 336 (6078): 233–7. Bibcode:2012Sci...336..233B. doi:10.1126/science.1215704. PMC 3547538. PMID 22422859.
^ Djuranovic S, Nahvi A, Green R (April 2012). "miRNA-mediated gene silencing by translational repression followed by mRNA deadenylation and decay". Science. 336 (6078): 237–40. Bibcode:2012Sci...336..237B. doi:10.1126/science.1215691. PMC 3971879. PMID 22499947.
^ Tan Y, Zhang B, Wu T, Skogerbø G, Zhu X, Guo X, He S, Chen R (February 2009). "Transcriptional inhibiton of Hoxd4 expression by miRNA-10a in human breast cancer cells". BMC Molecular Biology. 10 (1): 12. doi:10.1186/1471-2199-10-12. PMC 2680403. PMID 19232136.
^ Hawkins PG, Morris KV (March 2008). "RNA and transcriptional modulation of gene expression". Cell Cycle. 7 (5): 602–7. doi:10.4161/cc.7.5.5522. PMC 2877389. PMID 18256543.
^ Stark A, Brennecke J, Bushati N, Russell RB, Cohen SM (December 2005). "Animal MicroRNAs confer robustness to gene expression and have a significant impact on 3'UTR evolution". Cell. 123 (6): 1133–46. doi:10.1016/j.cell.2005.11.023. PMID 16337999.
^ Li LC (2008). "Small RNA-Mediated Gene Activation". In Morris KV. RNA and the Regulation of Gene Expression: A Hidden Layer of Complexity. Horizon Scientific Press. ISBN 978-1-904455-25-7.
^ Place RF, Li LC, Pookot D, Noonan EJ, Dahiya R (February 2008). "MicroRNA-373 induces expression of genes with complementary promoter sequences". Proceedings of the National Academy of Sciences of the United States of America. 105 (5): 1608–13. Bibcode:2008PNAS..105.1608P. doi:10.1073/pnas.0707594105. PMC 2234192. PMID 18227514.
^ Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP (August 2011). "A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language?". Cell. 146 (3): 353–8. doi:10.1016/j.cell.2011.07.014. PMC 3235919. PMID 21802130.
^ Axtell MJ, Bartel DP (June 2005). "Antiquity of microRNAs and their targets in land plants". The Plant Cell. 17 (6): 1658–73. doi:10.1105/tpc.105.032185. PMC 1143068. PMID 15849273.
^ Tanzer A, Stadler PF (May 2004). "Molecular evolution of a microRNA cluster". Journal of Molecular Biology. 339 (2): 327–35. CiteSeerX 10.1.1.194.1598. doi:10.1016/j.jmb.2004.03.065. PMID 15136036.
^ Chen K, Rajewsky N (February 2007). "The evolution of gene regulation by transcription factors and microRNAs". Nature Reviews. Genetics. 8 (2): 93–103. doi:10.1038/nrg1990. PMID 17230196.
^ Lee CT, Risom T, Strauss WM (April 2007). "Evolutionary conservation of microRNA regulatory circuits: an examination of microRNA gene complexity and conserved microRNA-target interactions through metazoan phylogeny". DNA and Cell Biology. 26 (4): 209–18. doi:10.1089/dna.2006.0545. PMID 17465887.
^ abcd Peterson KJ, Dietrich MR, McPeek MA (July 2009). "MicroRNAs and metazoan macroevolution: insights into canalization, complexity, and the Cambrian explosion". BioEssays. 31 (7): 736–47. doi:10.1002/bies.200900033. PMID 19472371.
^ Shabalina SA, Koonin EV (October 2008). "Origins and evolution of eukaryotic RNA interference". Trends in Ecology & Evolution. 23 (10): 578–87. doi:10.1016/j.tree.2008.06.005. PMC 2695246. PMID 18715673.
^ Axtell MJ, Westholm JO, Lai EC (2011). "Vive la différence: biogenesis and evolution of microRNAs in plants and animals". Genome Biology. 12 (4): 221. doi:10.1186/gb-2011-12-4-221. PMC 3218855. PMID 21554756.
^ ab Wheeler BM, Heimberg AM, Moy VN, Sperling EA, Holstein TW, Heber S, Peterson KJ (2009). "The deep evolution of metazoan microRNAs". Evolution & Development. 11 (1): 50–68. doi:10.1111/j.1525-142X.2008.00302.x. PMID 19196333.
^ Pashkovskiy PP, Ryazansky SS (June 2013). "Biogenesis, evolution, and functions of plant microRNAs". Biochemistry. Biokhimiia. 78 (6): 627–37. doi:10.1134/S0006297913060084. PMID 23980889.
^ ab Heimberg AM, Sempere LF, Moy VN, Donoghue PC, Peterson KJ (February 2008). "MicroRNAs and the advent of vertebrate morphological complexity". Proceedings of the National Academy of Sciences of the United States of America. 105 (8): 2946–50. Bibcode:2008PNAS..105.2946H. doi:10.1073/pnas.0712259105. PMC 2268565. PMID 18287013.
^ abc Nozawa M, Miura S, Nei M (July 2010). "Origins and evolution of microRNA genes in Drosophila species". Genome Biology and Evolution. 2: 180–9. doi:10.1093/gbe/evq009. PMC 2942034. PMID 20624724.
^ Allen E, Xie Z, Gustafson AM, Sung GH, Spatafora JW, Carrington JC (December 2004). "Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana". Nature Genetics. 36 (12): 1282–90. doi:10.1038/ng1478. PMID 15565108.
^ Warthmann N, Das S, Lanz C, Weigel D (May 2008). "Comparative analysis of the MIR319a microRNA locus in Arabidopsis and related Brassicaceae". Molecular Biology and Evolution. 25 (5): 892–902. doi:10.1093/molbev/msn029. PMID 18296705.
^ Fahlgren N, Jogdeo S, Kasschau KD, Sullivan CM, Chapman EJ, Laubinger S, Smith LM, Dasenko M, Givan SA, Weigel D, Carrington JC (April 2010). "MicroRNA gene evolution in Arabidopsis lyrata and Arabidopsis thaliana". The Plant Cell. 22 (4): 1074–89. doi:10.1105/tpc.110.073999. PMC 2879733. PMID 20407027.
^ Caravas J, Friedrich M (June 2010). "Of mites and millipedes: recent progress in resolving the base of the arthropod tree". BioEssays. 32 (6): 488–95. doi:10.1002/bies.201000005. PMID 20486135.
^ Kenny NJ, Namigai EK, Marlétaz F, Hui JH, Shimeld SM (December 2015). "Draft genome assemblies and predicted microRNA complements of the intertidal lophotrochozoans Patella vulgata (Mollusca, Patellogastropoda) and Spirobranchus (Pomatoceros) lamarcki (Annelida, Serpulida)". Marine Genomics. 24 Pt 2 (2): 139–46. doi:10.1016/j.margen.2015.07.004. PMID 26319627.
^ Cock JM, Sterck L, Rouzé P, Scornet D, Allen AE, Amoutzias G, et al. (June 2010). "The Ectocarpus genome and the independent evolution of multicellularity in brown algae". Nature. 465 (7298): 617–21. Bibcode:2010Natur.465..617C. doi:10.1038/nature09016. PMID 20520714.
^ Cuperus JT, Fahlgren N, Carrington JC (February 2011). "Evolution and functional diversification of MIRNA genes". The Plant Cell. 23 (2): 431–42. doi:10.1105/tpc.110.082784. PMC 3077775. PMID 21317375.
^ Ryan JF, Pang K, Schnitzler CE, Nguyen AD, Moreland RT, Simmons DK, Koch BJ, Francis WR, Havlak P, Smith SA, Putnam NH, Haddock SH, Dunn CW, Wolfsberg TG, Mullikin JC, Martindale MQ, Baxevanis AD (December 2013). "The genome of the ctenophore Mnemiopsis leidyi and its implications for cell type evolution". Science. 342 (6164): 1242592. doi:10.1126/science.1242592. PMC 3920664. PMID 24337300.
^ Maxwell EK, Ryan JF, Schnitzler CE, Browne WE, Baxevanis AD (December 2012). "MicroRNAs and essential components of the microRNA processing machinery are not encoded in the genome of the ctenophore Mnemiopsis leidyi". BMC Genomics. 13 (1): 714. doi:10.1186/1471-2164-13-714. PMC 3563456. PMID 23256903.
^ Dimond PF (15 March 2010). "miRNAs' Therapeutic Potential". Genetic Engineering & Biotechnology News. 30 (6): 1. Archived from the original on 10 July 2010. Retrieved 10 July 2010.
^ Tjaden B, Goodwin SS, Opdyke JA, Guillier M, Fu DX, Gottesman S, Storz G (2006). "Target prediction for small, noncoding RNAs in bacteria". Nucleic Acids Research. 34 (9): 2791–802. doi:10.1093/nar/gkl356. PMC 1464411. PMID 16717284.
^ Liu CG, Calin GA, Volinia S, Croce CM (2008). "MicroRNA expression profiling using microarrays". Nature Protocols. 3 (4): 563–78. doi:10.1038/nprot.2008.14. PMID 18388938.
^ Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH, Nguyen JT, Barbisin M, Xu NL, Mahuvakar VR, Andersen MR, Lao KQ, Livak KJ, Guegler KJ (November 2005). "Real-time quantification of microRNAs by stem-loop RT-PCR". Nucleic Acids Research. 33 (20): e179. doi:10.1093/nar/gni178. PMC 1292995. PMID 16314309.
^ Shingara J, Keiger K, Shelton J, Laosinchai-Wolf W, Powers P, Conrad R, Brown D, Labourier E (September 2005). "An optimized isolation and labeling platform for accurate microRNA expression profiling". RNA. 11 (9): 1461–70. doi:10.1261/rna.2610405. PMC 1370829. PMID 16043497.
^ Buermans HP, Ariyurek Y, van Ommen G, den Dunnen JT, 't Hoen PA (December 2010). "New methods for next generation sequencing based microRNA expression profiling". BMC Genomics. 11: 716. doi:10.1186/1471-2164-11-716. PMC 3022920. PMID 21171994.
^ Kloosterman WP, Wienholds E, Ketting RF, Plasterk RH (2004). "Substrate requirements for let-7 function in the developing zebrafish embryo". Nucleic Acids Research. 32 (21): 6284–91. doi:10.1093/nar/gkh968. PMC 535676. PMID 15585662.
^ Flynt AS, Li N, Thatcher EJ, Solnica-Krezel L, Patton JG (February 2007). "Zebrafish miR-214 modulates Hedgehog signaling to specify muscle cell fate". Nature Genetics. 39 (2): 259–63. doi:10.1038/ng1953. PMC 3982799. PMID 17220889.
^ Meister G, Landthaler M, Dorsett Y, Tuschl T (March 2004). "Sequence-specific inhibition of microRNA- and siRNA-induced RNA silencing". RNA. 10 (3): 544–50. doi:10.1261/rna.5235104. PMC 1370948. PMID 14970398.
^ Kloosterman WP, Lagendijk AK, Ketting RF, Moulton JD, Plasterk RH (August 2007). "Targeted inhibition of miRNA maturation with morpholinos reveals a role for miR-375 in pancreatic islet development". PLoS Biology. 5 (8): e203. doi:10.1371/journal.pbio.0050203. PMC 1925136. PMID 17676975.
^ Gebert LF, Rebhan MA, Crivelli SE, Denzler R, Stoffel M, Hall J (January 2014). "Miravirsen (SPC3649) can inhibit the biogenesis of miR-122". Nucleic Acids Research. 42 (1): 609–21. doi:10.1093/nar/gkt852. PMC 3874169. PMID 24068553.
^ Choi WY, Giraldez AJ, Schier AF (October 2007). "Target protectors reveal dampening and balancing of Nodal agonist and antagonist by miR-430". Science. 318 (5848): 271–4. Bibcode:2007Sci...318..271C. doi:10.1126/science.1147535. PMID 17761850.
^ You Y, Moreira BG, Behlke MA, Owczarzy R (May 2006). "Design of LNA probes that improve mismatch discrimination". Nucleic Acids Research. 34 (8): e60. doi:10.1093/nar/gkl175. PMC 1456327. PMID 16670427.
^ Lagendijk AK, Moulton JD, Bakkers J (June 2012). "Revealing details: whole mount microRNA in situ hybridization protocol for zebrafish embryos and adult tissues". Biology Open. 1 (6): 566–9. doi:10.1242/bio.2012810. PMC 3509442. PMID 23213449.
^ Kaur H, Arora A, Wengel J, Maiti S (June 2006). "Thermodynamic, counterion, and hydration effects for the incorporation of locked nucleic acid nucleotides into DNA duplexes". Biochemistry. 45 (23): 7347–55. doi:10.1021/bi060307w. PMID 16752924.
^ Nielsen JA, Lau P, Maric D, Barker JL, Hudson LD (August 2009). "Integrating microRNA and mRNA expression profiles of neuronal progenitors to identify regulatory networks underlying the onset of cortical neurogenesis". BMC Neuroscience. 10: 98. doi:10.1186/1471-2202-10-98. PMC 2736963. PMID 19689821.
^ Gupta A, Nagilla P, Le HS, Bunney C, Zych C, Thalamuthu A, Bar-Joseph Z, Mathavan S, Ayyavoo V (2011). Mammano F, ed. "Comparative expression profile of miRNA and mRNA in primary peripheral blood mononuclear cells infected with human immunodeficiency virus (HIV-1)". PLOS One. 6 (7): e22730. Bibcode:2011PLoSO...622730G. doi:10.1371/journal.pone.0022730. PMC 3145673. PMID 21829495.
^ Grimson A, Farh KK, Johnston WK, Garrett-Engele P, Lim LP, Bartel DP (July 2007). "MicroRNA targeting specificity in mammals: determinants beyond seed pairing". Molecular Cell. 27 (1): 91–105. doi:10.1016/j.molcel.2007.06.017. PMC 3800283. PMID 17612493.
^ Griffiths-Jones S, Saini HK, van Dongen S, Enright AJ (January 2008). "miRBase: tools for microRNA genomics". Nucleic Acids Research. 36 (Database issue): D154–8. doi:10.1093/nar/gkm952. PMC 2238936. PMID 17991681.
^ Nam S, Li M, Choi K, Balch C, Kim S, Nephew KP (July 2009). "MicroRNA and mRNA integrated analysis (MMIA): a web tool for examining biological functions of microRNA expression". Nucleic Acids Research. 37 (Web Server issue): W356–62. doi:10.1093/nar/gkp294. PMC 2703907. PMID 19420067.
^ Artmann S, Jung K, Bleckmann A, Beissbarth T (2012). Provero P, ed. "Detection of simultaneous group effects in microRNA expression and related target gene sets". PLOS One. 7 (6): e38365. Bibcode:2012PLoSO...738365A. doi:10.1371/journal.pone.0038365. PMC 3378551. PMID 22723856.
^ Jiang Q, Wang Y, Hao Y, Juan L, Teng M, Zhang X, Li M, Wang G, Liu Y (January 2009). "miR2Disease: a manually curated database for microRNA deregulation in human disease". Nucleic Acids Research. 37. 37 (Database issue): D98–104. doi:10.1093/nar/gkn714. PMC 2686559. PMID 18927107.
^ Mencía A, Modamio-Høybjør S, Redshaw N, Morín M, Mayo-Merino F, Olavarrieta L, Aguirre LA, del Castillo I, Steel KP, Dalmay T, Moreno F, Moreno-Pelayo MA (May 2009). "Mutations in the seed region of human miR-96 are responsible for nonsyndromic progressive hearing loss". Nature Genetics. 41 (5): 609–13. doi:10.1038/ng.355. PMID 19363479.
^ Hughes AE, Bradley DT, Campbell M, Lechner J, Dash DP, Simpson DA, Willoughby CE (November 2011). "Mutation altering the miR-184 seed region causes familial keratoconus with cataract". American Journal of Human Genetics. 89 (5): 628–33. doi:10.1016/j.ajhg.2011.09.014. PMC 3213395. PMID 21996275.
^ de Pontual L, Yao E, Callier P, Faivre L, Drouin V, Cariou S, Van Haeringen A, Geneviève D, Goldenberg A, Oufadem M, Manouvrier S, Munnich A, Vidigal JA, Vekemans M, Lyonnet S, Henrion-Caude A, Ventura A, Amiel J (September 2011). "Germline deletion of the miR-17∼92 cluster causes skeletal and growth defects in humans". Nature Genetics. 43 (10): 1026–30. doi:10.1038/ng.915. PMC 3184212. PMID 21892160.
^ Võsa U, Vooder T, Kolde R, Fischer K, Välk K, Tõnisson N, Roosipuu R, Vilo J, Metspalu A, Annilo T (October 2011). "Identification of miR-374a as a prognostic marker for survival in patients with early-stage nonsmall cell lung cancer". Genes, Chromosomes & Cancer. 50 (10): 812–22. doi:10.1002/gcc.20902. PMID 21748820.
^ Akçakaya P, Ekelund S, Kolosenko I, Caramuta S, Ozata DM, Xie H, Lindforss U, Olivecrona H, Lui WO (August 2011). "miR-185 and miR-133b deregulation is associated with overall survival and metastasis in colorectal cancer". International Journal of Oncology. 39 (2): 311–8. doi:10.3892/ijo.2011.1043. PMID 21573504.
^ ab Eyking A, Reis H, Frank M, Gerken G, Schmid KW, Cario E (2016). "MiR-205 and MiR-373 Are Associated with Aggressive Human Mucinous Colorectal Cancer". PLOS One. 11 (6): e0156871. Bibcode:2016PLoSO..1156871E. doi:10.1371/journal.pone.0156871. PMC 4894642. PMID 27271572.
^ MicroRNA-21 promotes hepatocellular carcinoma HepG2 cell proliferation through repression of mitogen-activated protein kinase-kinase 3. Guangxian Xu et al, 2013
^ Jones K, Nourse JP, Keane C, Bhatnagar A, Gandhi MK (January 2014). "Plasma microRNA are disease response biomarkers in classical Hodgkin lymphoma". Clinical Cancer Research. 20 (1): 253–64. doi:10.1158/1078-0432.CCR-13-1024. PMID 24222179.
^ Hosseinahli N, Aghapour M, Duijf PH, Baradaran B (August 2018). "Treating cancer with microRNA replacement therapy: A literature review". Journal of Cellular Physiology. 233 (8): 5574–5588. doi:10.1002/jcp.26514. PMID 29521426.
^ Liu G, Sun Y, Ji P, Li X, Cogdell D, Yang D, Parker Kerrigan BC, Shmulevich I, Chen K, Sood AK, Xue F, Zhang W (July 2014). "MiR-506 suppresses proliferation and induces senescence by directly targeting the CDK4/6-FOXM1 axis in ovarian cancer". The Journal of Pathology. 233 (3): 308–18. doi:10.1002/path.4348. PMC 4144705. PMID 24604117.
^ Wen SY, Lin Y, Yu YQ, Cao SJ, Zhang R, Yang XM, Li J, Zhang YL, Wang YH, Ma MZ, Sun WW, Lou XL, Wang JH, Teng YC, Zhang ZG (February 2015). "miR-506 acts as a tumor suppressor by directly targeting the hedgehog pathway transcription factor Gli3 in human cervical cancer". Oncogene. 34 (6): 717–25. doi:10.1038/onc.2014.9. PMID 24608427.
^ Bernstein C, Prasad AR, Nfonsam V, Bernstei H (2013). "DNA Damage, DNA Repair and Cancer". New Research Directions in DNA Repair. pp. 413–65. doi:10.5772/53919. ISBN 978-953-51-1114-6.
^ O'Hagan HM, Mohammad HP, Baylin SB (August 2008). "Double strand breaks can initiate gene silencing and SIRT1-dependent onset of DNA methylation in an exogenous promoter CpG island". PLoS Genetics. 4 (8): e1000155. doi:10.1371/journal.pgen.1000155. PMC 2491723. PMID 18704159.
^ Cuozzo C, Porcellini A, Angrisano T, Morano A, Lee B, Di Pardo A, Messina S, Iuliano R, Fusco A, Santillo MR, Muller MT, Chiariotti L, Gottesman ME, Avvedimento EV (July 2007). "DNA damage, homology-directed repair, and DNA methylation". PLoS Genetics. 3 (7): e110. doi:10.1371/journal.pgen.0030110. PMC 1913100. PMID 17616978.
^ Jasperson KW, Tuohy TM, Neklason DW, Burt RW (June 2010). "Hereditary and familial colon cancer". Gastroenterology. 138 (6): 2044–58. doi:10.1053/j.gastro.2010.01.054. PMC 3057468. PMID 20420945.
^ Truninger K, Menigatti M, Luz J, Russell A, Haider R, Gebbers JO, Bannwart F, Yurtsever H, Neuweiler J, Riehle HM, Cattaruzza MS, Heinimann K, Schär P, Jiricny J, Marra G (May 2005). "Immunohistochemical analysis reveals high frequency of PMS2 defects in colorectal cancer". Gastroenterology. 128 (5): 1160–71. doi:10.1053/j.gastro.2005.01.056. PMID 15887099.
^ Valeri N, Gasparini P, Fabbri M, Braconi C, Veronese A, Lovat F, Adair B, Vannini I, Fanini F, Bottoni A, Costinean S, Sandhu SK, Nuovo GJ, Alder H, Gafa R, Calore F, Ferracin M, Lanza G, Volinia S, Negrini M, McIlhatton MA, Amadori D, Fishel R, Croce CM (April 2010). "Modulation of mismatch repair and genomic stability by miR-155". Proceedings of the National Academy of Sciences of the United States of America. 107 (15): 6982–7. Bibcode:2010PNAS..107.6982V. doi:10.1073/pnas.1002472107. PMC 2872463. PMID 20351277.
^ abc Zhang W, Zhang J, Hoadley K, Kushwaha D, Ramakrishnan V, Li S, Kang C, You Y, Jiang C, Song SW, Jiang T, Chen CC (June 2012). "miR-181d: a predictive glioblastoma biomarker that downregulates MGMT expression". Neuro-Oncology. 14 (6): 712–9. doi:10.1093/neuonc/nos089. PMC 3367855. PMID 22570426.
^ Spiegl-Kreinecker S, Pirker C, Filipits M, Lötsch D, Buchroithner J, Pichler J, Silye R, Weis S, Micksche M, Fischer J, Berger W (January 2010). "O6-Methylguanine DNA methyltransferase protein expression in tumor cells predicts outcome of temozolomide therapy in glioblastoma patients". Neuro-Oncology. 12 (1): 28–36. doi:10.1093/neuonc/nop003. PMC 2940563. PMID 20150365.
^ Sgarra R, Rustighi A, Tessari MA, Di Bernardo J, Altamura S, Fusco A, Manfioletti G, Giancotti V (September 2004). "Nuclear phosphoproteins HMGA and their relationship with chromatin structure and cancer". FEBS Letters. 574 (1–3): 1–8. doi:10.1016/j.febslet.2004.08.013. PMID 15358530.
^ Xu Y, Sumter TF, Bhattacharya R, Tesfaye A, Fuchs EJ, Wood LJ, Huso DL, Resar LM (May 2004). "The HMG-I oncogene causes highly penetrant, aggressive lymphoid malignancy in transgenic mice and is overexpressed in human leukemia". Cancer Research. 64 (10): 3371–5. doi:10.1158/0008-5472.CAN-04-0044. PMID 15150086.
^ Borrmann L, Schwanbeck R, Heyduk T, Seebeck B, Rogalla P, Bullerdiek J, Wisniewski JR (December 2003). "High mobility group A2 protein and its derivatives bind a specific region of the promoter of DNA repair gene ERCC1 and modulate its activity". Nucleic Acids Research. 31 (23): 6841–51. doi:10.1093/nar/gkg884. PMC 290254. PMID 14627817.
^ Facista A, Nguyen H, Lewis C, Prasad AR, Ramsey L, Zaitlin B, Nfonsam V, Krouse RS, Bernstein H, Payne CM, Stern S, Oatman N, Banerjee B, Bernstein C (April 2012). "Deficient expression of DNA repair enzymes in early progression to sporadic colon cancer". Genome Integrity. 3 (1): 3. doi:10.1186/2041-9414-3-3. PMC 3351028. PMID 22494821.
^ Chen JF, Murchison EP, Tang R, Callis TE, Tatsuguchi M, Deng Z, Rojas M, Hammond SM, Schneider MD, Selzman CH, Meissner G, Patterson C, Hannon GJ, Wang DZ (February 2008). "Targeted deletion of Dicer in the heart leads to dilated cardiomyopathy and heart failure". Proceedings of the National Academy of Sciences of the United States of America. 105 (6): 2111–6. Bibcode:2008PNAS..105.2111C. doi:10.1073/pnas.0710228105. PMC 2542870. PMID 18256189.
^ ab Zhao Y, Ransom JF, Li A, Vedantham V, von Drehle M, Muth AN, Tsuchihashi T, McManus MT, Schwartz RJ, Srivastava D (April 2007). "Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2". Cell. 129 (2): 303–17. doi:10.1016/j.cell.2007.03.030. PMID 17397913.
^ Thum T, Galuppo P, Wolf C, Fiedler J, Kneitz S, van Laake LW, Doevendans PA, Mummery CL, Borlak J, Haverich A, Gross C, Engelhardt S, Ertl G, Bauersachs J (July 2007). "MicroRNAs in the human heart: a clue to fetal gene reprogramming in heart failure". Circulation. 116 (3): 258–67. doi:10.1161/CIRCULATIONAHA.107.687947. PMID 17606841.
^ van Rooij E, Sutherland LB, Liu N, Williams AH, McAnally J, Gerard RD, Richardson JA, Olson EN (November 2006). "A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure". Proceedings of the National Academy of Sciences of the United States of America. 103 (48): 18255–60. Bibcode:2006PNAS..10318255V. doi:10.1073/pnas.0608791103. PMC 1838739. PMID 17108080.
^ Tatsuguchi M, Seok HY, Callis TE, Thomson JM, Chen JF, Newman M, Rojas M, Hammond SM, Wang DZ (June 2007). "Expression of microRNAs is dynamically regulated during cardiomyocyte hypertrophy". Journal of Molecular and Cellular Cardiology. 42 (6): 1137–41. doi:10.1016/j.yjmcc.2007.04.004. PMC 1934409. PMID 17498736.
^ Zhao Y, Samal E, Srivastava D (July 2005). "Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis". Nature. 436 (7048): 214–20. Bibcode:2005Natur.436..214Z. doi:10.1038/nature03817. PMID 15951802.
^ Xiao J, Luo X, Lin H, Zhang Y, Lu Y, Wang N, Zhang Y, Yang B, Wang Z (April 2007). "MicroRNA miR-133 represses HERG K+ channel expression contributing to QT prolongation in diabetic hearts". The Journal of Biological Chemistry. 282 (17): 12363–7. doi:10.1074/jbc.C700015200. PMC 3151106. PMID 17344217.
^ Yang B, Lin H, Xiao J, Lu Y, Luo X, Li B, Zhang Y, Xu C, Bai Y, Wang H, Chen G, Wang Z (April 2007). "The muscle-specific microRNA miR-1 regulates cardiac arrhythmogenic potential by targeting GJA1 and KCNJ2". Nature Medicine. 13 (4): 486–91. doi:10.1038/nm1569. PMID 17401374.
^ Carè A, Catalucci D, Felicetti F, Bonci D, Addario A, Gallo P, Bang ML, Segnalini P, Gu Y, Dalton ND, Elia L, Latronico MV, Høydal M, Autore C, Russo MA, Dorn GW, Ellingsen O, Ruiz-Lozano P, Peterson KL, Croce CM, Peschle C, Condorelli G (May 2007). "MicroRNA-133 controls cardiac hypertrophy". Nature Medicine. 13 (5): 613–8. doi:10.1038/nm1582. PMID 17468766.
^ van Rooij E, Sutherland LB, Qi X, Richardson JA, Hill J, Olson EN (April 2007). "Control of stress-dependent cardiac growth and gene expression by a microRNA". Science. 316 (5824): 575–9. Bibcode:2007Sci...316..575V. doi:10.1126/science.1139089. PMID 17379774.
^ Keller T, Boeckel JN, Groß S, Klotsche J, Palapies L, Leistner D, et al. (July 2017). "Improved risk stratification in prevention by use of a panel of selected circulating microRNAs". Scientific Reports. 7 (1): 4511. Bibcode:2017NatSR...7.4511K. doi:10.1038/s41598-017-04040-w. PMC 5495799. PMID 28674420.
^ Wagschal A, Najafi-Shoushtari SH, Wang L, Goedeke L, Sinha S, deLemos AS, et al. (November 2015). "Genome-wide identification of microRNAs regulating cholesterol and triglyceride homeostasis". Nature Medicine. 21 (11): 1290–7. doi:10.1038/nm.3980. PMC 4993048. PMID 26501192.
^ Insull W (January 2009). "The pathology of atherosclerosis: plaque development and plaque responses to medical treatment". The American Journal of Medicine. 122 (1 Suppl): S3–S14. doi:10.1016/j.amjmed.2008.10.013. PMID 19110086.
^ abcdefghijklmnopq Son DJ, Kumar S, Takabe W, Kim CW, Ni CW, Alberts-Grill N, Jang IH, Kim S, Kim W, Won Kang S, Baker AH, Woong Seo J, Ferrara KW, Jo H (2013). "The atypical mechanosensitive microRNA-712 derived from pre-ribosomal RNA induces endothelial inflammation and atherosclerosis". Nature Communications. 4: 3000. Bibcode:2013NatCo...4E3000S. doi:10.1038/ncomms4000. PMC 3923891. PMID 24346612.
^ ab Basu R, Fan D, Kandalam V, Lee J, Das SK, Wang X, Baldwin TA, Oudit GY, Kassiri Z (December 2012). "Loss of Timp3 gene leads to abdominal aortic aneurysm formation in response to angiotensin II". The Journal of Biological Chemistry. 287 (53): 44083–96. doi:10.1074/jbc.M112.425652. PMC 3531724. PMID 23144462.
^ Libby P (2002). "Inflammation in atherosclerosis". Nature. 420 (6917): 868–74. Bibcode:2002Natur.420..868L. doi:10.1038/nature01323. PMID 12490960.
^ ab Phua YL, Chu JY, Marrone AK, Bodnar AJ, Sims-Lucas S, Ho J (October 2015). "Renal stromal miRNAs are required for normal nephrogenesis and glomerular mesangial survival". Physiological Reports. 3 (10): e12537. doi:10.14814/phy2.12537. PMC 4632944. PMID 26438731.
^ Maes OC, Chertkow HM, Wang E, Schipper HM (May 2009). "MicroRNA: Implications for Alzheimer Disease and other Human CNS Disorders". Current Genomics. 10 (3): 154–68. doi:10.2174/138920209788185252. PMC 2705849. PMID 19881909.
^ Amin ND, Bai G, Klug JR, Bonanomi D, Pankratz MT, Gifford WD, Hinckley CA, Sternfeld MJ, Driscoll SP, Dominguez B, Lee KF, Jin X, Pfaff SL (December 2015). "Loss of motoneuron-specific microRNA-218 causes systemic neuromuscular failure". Science. 350 (6267): 1525–9. Bibcode:2015Sci...350.1525A. doi:10.1126/science.aad2509. PMC 4913787. PMID 26680198.
^ Schratt G (December 2009). "microRNAs at the synapse". Nature Reviews. Neuroscience. 10 (12): 842–9. doi:10.1038/nrn2763. PMID 19888283.
^ Hommers LG, Domschke K, Deckert J (January 2015). "Heterogeneity and individuality: microRNAs in mental disorders". Journal of Neural Transmission. 122 (1): 79–97. doi:10.1007/s00702-014-1338-4. PMID 25395183.
^ Feng J, Sun G, Yan J, Noltner K, Li W, Buzin CH, Longmate J, Heston LL, Rossi J, Sommer SS (July 2009). Reif A, ed. "Evidence for X-chromosomal schizophrenia associated with microRNA alterations". PLOS One. 4 (7): e6121. Bibcode:2009PLoSO...4.6121F. doi:10.1371/journal.pone.0006121. PMC 2699475. PMID 19568434.
^ Beveridge NJ, Gardiner E, Carroll AP, Tooney PA, Cairns MJ (December 2010). "Schizophrenia is associated with an increase in cortical microRNA biogenesis". Molecular Psychiatry. 15 (12): 1176–89. doi:10.1038/mp.2009.84. PMC 2990188. PMID 19721432.
^ abc Lewohl JM, Nunez YO, Dodd PR, Tiwari GR, Harris RA, Mayfield RD (November 2011). "Up-regulation of microRNAs in brain of human alcoholics". Alcoholism, Clinical and Experimental Research. 35 (11): 1928–37. doi:10.1111/j.1530-0277.2011.01544.x. PMC 3170679. PMID 21651580.
^ Tapocik JD, Solomon M, Flanigan M, Meinhardt M, Barbier E, Schank JR, Schwandt M, Sommer WH, Heilig M (June 2013). "Coordinated dysregulation of mRNAs and microRNAs in the rat medial prefrontal cortex following a history of alcohol dependence". The Pharmacogenomics Journal. 13 (3): 286–96. doi:10.1038/tpj.2012.17. PMC 3546132. PMID 22614244.
^ Gorini G, Nunez YO, Mayfield RD (2013). "Integration of miRNA and protein profiling reveals coordinated neuroadaptations in the alcohol-dependent mouse brain". PLOS One. 8 (12): e82565. Bibcode:2013PLoSO...882565G. doi:10.1371/journal.pone.0082565. PMC 3865091. PMID 24358208.
^ Tapocik JD, Barbier E, Flanigan M, Solomon M, Pincus A, Pilling A, Sun H, Schank JR, King C, Heilig M (March 2014). "microRNA-206 in rat medial prefrontal cortex regulates BDNF expression and alcohol drinking". The Journal of Neuroscience. 34 (13): 4581–8. doi:10.1523/JNEUROSCI.0445-14.2014. PMC 3965783. PMID 24672003.
^ Lippai D, Bala S, Csak T, Kurt-Jones EA, Szabo G (2013). "Chronic alcohol-induced microRNA-155 contributes to neuroinflammation in a TLR4-dependent manner in mice". PLOS One. 8 (8): e70945. Bibcode:2013PLoSO...870945L. doi:10.1371/journal.pone.0070945. PMC 3739772. PMID 23951048.
^ ab Li J, Li J, Liu X, Qin S, Guan Y, Liu Y, Cheng Y, Chen X, Li W, Wang S, Xiong M, Kuzhikandathil EV, Ye JH, Zhang C (September 2013). "MicroRNA expression profile and functional analysis reveal that miR-382 is a critical novel gene of alcohol addiction". EMBO Molecular Medicine. 5 (9): 1402–14. doi:10.1002/emmm.201201900. PMC 3799494. PMID 23873704.
^ Romao JM, Jin W, Dodson MV, Hausman GJ, Moore SS, Guan LL (September 2011). "MicroRNA regulation in mammalian adipogenesis". Experimental Biology and Medicine. 236 (9): 997–1004. doi:10.1258/ebm.2011.011101. PMID 21844119.
^ Skårn M, Namløs HM, Noordhuis P, Wang MY, Meza-Zepeda LA, Myklebost O (April 2012). "Adipocyte differentiation of human bone marrow-derived stromal cells is modulated by microRNA-155, microRNA-221, and microRNA-222". Stem Cells and Development. 21 (6): 873–83. doi:10.1089/scd.2010.0503. PMID 21756067.
^ Zuo Y, Qiang L, Farmer SR (March 2006). "Activation of CCAAT/enhancer-binding protein (C/EBP) alpha expression by C/EBP beta during adipogenesis requires a peroxisome proliferator-activated receptor-gamma-associated repression of HDAC1 at the C/ebp alpha gene promoter". The Journal of Biological Chemistry. 281 (12): 7960–7. doi:10.1074/jbc.M510682200. PMID 16431920.
^ Jun-Hao ET, Gupta RR, Shyh-Chang N (March 2016). "Lin28 and let-7 in the Metabolic Physiology of Aging". Trends in Endocrinology and Metabolism. 27 (3): 132–141. doi:10.1016/j.tem.2015.12.006. PMID 26811207.
^ Zhu H, Shyh-Chang N, Segrè AV, Shinoda G, Shah SP, Einhorn WS, Takeuchi A, Engreitz JM, Hagan JP, Kharas MG, Urbach A, Thornton JE, Triboulet R, Gregory RI, Altshuler D & Daley GQ (September 2011). "The Lin28/let-7 axis regulates glucose metabolism". Cell. 147 (1): 81–94. doi:10.1016/j.cell.2011.08.033. PMC 3353524. PMID 21962509.
^ Frost RJ, Olson EN (December 2011). "Control of glucose homeostasis and insulin sensitivity by the Let-7 family of microRNAs". Proceedings of the National Academy of Sciences of the United States of America. 108 (52): 21075–80. Bibcode:2011PNAS..10821075F. doi:10.1073/pnas.1118922109. PMC 3248488. PMID 22160727.
^ Teruel-Montoya R, Rosendaal FR, Martínez C (February 2015). "MicroRNAs in hemostasis". J Thromb Haemost. 13 (2): 170–181. PMID 25400249.
^ ab Nourse J, Braun J, Lackner K, Hüttelmaier S, Danckwardt S (September 2018). "Large-scale identification of functional microRNA targeting reveals cooperative regulation of the hemostatic system". J Thromb Haemost. doi:10.1111/jth.14290. PMID 30207063.
^ Pheasant M, Mattick JS (September 2007). "Raising the estimate of functional human sequences". Genome Research. 17 (9): 1245–53. doi:10.1101/gr.6406307. PMID 17690206.
^ Bertone P, Stolc V, Royce TE, Rozowsky JS, Urban AE, Zhu X, Rinn JL, Tongprasit W, Samanta M, Weissman S, Gerstein M, Snyder M (December 2004). "Global identification of human transcribed sequences with genome tiling arrays". Science. 306 (5705): 2242–6. Bibcode:2004Sci...306.2242B. doi:10.1126/science.1103388. PMID 15539566.
^ Ota T, Suzuki Y, Nishikawa T, Otsuki T, Sugiyama T, Irie R, et al. (January 2004). "Complete sequencing and characterization of 21,243 full-length human cDNAs". Nature Genetics. 36 (1): 40–5. doi:10.1038/ng1285. PMID 14702039.
^ Kuhn DE, Martin MM, Feldman DS, Terry AV, Nuovo GJ, Elton TS (January 2008). "Experimental validation of miRNA targets". Methods. 44 (1): 47–54. doi:10.1016/j.ymeth.2007.09.005. PMC 2237914. PMID 18158132.
^ Hüttenhofer A, Schattner P, Polacek N (May 2005). "Non-coding RNAs: hope or hype?". Trends in Genetics. 21 (5): 289–97. doi:10.1016/j.tig.2005.03.007. PMID 15851066.
^ Qureshi A, Thakur N, Monga I, Thakur A, Kumar M (2014-01-01). "VIRmiRNA: a comprehensive resource for experimentally validated viral miRNAs and their targets". Database. 2014: bau103. doi:10.1093/database/bau103. PMC 4224276. PMID 25380780.
^ Kumar M. "VIRmiRNA". Resource for experimental viral miRNA and their targets. Bioinformatics center, CSIR-IMTECH.
^ Tuddenham L, Jung JS, Chane-Woon-Ming B, Dölken L, Pfeffer S (February 2012). "Small RNA deep sequencing identifies microRNAs and other small noncoding RNAs from human herpesvirus 6B". Journal of Virology. 86 (3): 1638–49. doi:10.1128/JVI.05911-11. PMC 3264354. PMID 22114334.
^ Zheng H, Fu R, Wang JT, Liu Q, Chen H, Jiang SW (April 2013). "Advances in the techniques for the prediction of microRNA targets". International Journal of Molecular Sciences. 14 (4): 8179–87. doi:10.3390/ijms14048179. PMC 3645737. PMID 23591837.
^ Agarwal V, Bell GW, Nam JW, Bartel DP (August 2015). "Predicting effective microRNA target sites in mammalian mRNAs". eLife. 4: e05005. doi:10.7554/eLife.05005. PMC 4532895. PMID 26267216.
Further reading
.mw-parser-output .refbeginfont-size:90%;margin-bottom:0.5em.mw-parser-output .refbegin-hanging-indents>ullist-style-type:none;margin-left:0.mw-parser-output .refbegin-hanging-indents>ul>li,.mw-parser-output .refbegin-hanging-indents>dl>ddmargin-left:0;padding-left:3.2em;text-indent:-3.2em;list-style:none.mw-parser-output .refbegin-100font-size:100%
miRNA definition and classification: Ambros V, Bartel B, Bartel DP, Burge CB, Carrington JC, Chen X, Dreyfuss G, Eddy SR, Griffiths-Jones S, Marshall M, Matzke M, Ruvkun G, Tuschl T (March 2003). "A uniform system for microRNA annotation". RNA. 9 (3): 277–9. doi:10.1261/rna.2183803. PMC 1370393. PMID 12592000.
Science review of small RNA: Baulcombe D (September 2002). "DNA events. An RNA microcosm". Science. 297 (5589): 2002–3. doi:10.1126/science.1077906. PMID 12242426.
Discovery of lin-4, the first miRNA to be discovered: Lee RC, Feinbaum RL, Ambros V (December 1993). "The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14". Cell. 75 (5): 843–54. doi:10.1016/0092-8674(93)90529-Y. PMID 8252621.
External links
| Look up microRNA or miRNA in Wiktionary, the free dictionary. |
- The miRBase database
miRTarBase, the experimentally validated microRNA-target interactions database.
miR2Disease, a manually curated database documenting known relationships between miRNA dysregulation and human disease.
semirna, Web application to search for microRNAs in a plant genome.
ONCO.IO: Integrative resource for microRNA and transcription factors analysis in cancer.
MirOB: MicroRNA targets database and data analysis and dataviz tool.
ChIPBase database: An open access database for decoding the transcription factors that were involved in or affected the transcription of microRNAs from ChIP-seq data.
An animated video of the microRNA biogenesis process.
miRNA modulation reagents to enable up-regulation or suppression of endogenous mature microRNA function


